

Direct role for the RNA polymerase domain of T7 primase in primer delivery
- PMID: 20439755
- PMCID: PMC2889121
- DOI: 10.1073/pnas.1004220107
Direct role for the RNA polymerase domain of T7 primase in primer delivery
Abstract
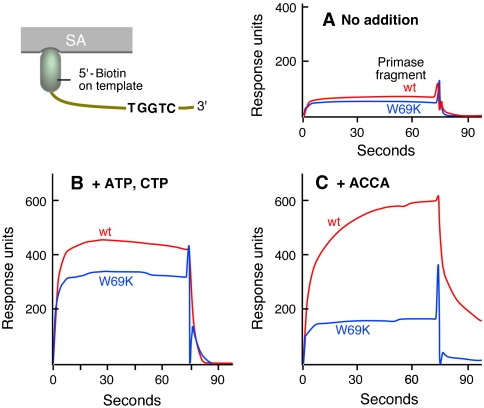
Gene 4 protein (gp4) encoded by bacteriophage T7 contains a C-terminal helicase and an N-terminal primase domain. After synthesis of tetraribonucleotides, gp4 must transfer them to the polymerase for use as primers to initiate DNA synthesis. In vivo gp4 exists in two molecular weight forms, a 56-kDa form and the full-length 63-kDa form. The 56-kDa gp4 lacks the N-terminal Cys(4) zinc-binding motif important in the recognition of primase sites in DNA. The 56-kDa gp4 is defective in primer synthesis but delivers a wider range of primers to initiate DNA synthesis compared to the 63-kDa gp4. Suppressors exist that enable the 56-kDa gp4 to support the growth of T7 phage lacking gene 4 (T7Delta4). We have identified 56-kDa DNA primases defective in primer delivery by screening for their ability to support growth of T7Delta4 phage in the presence of this suppressor. Trp69 is critical for primer delivery. Replacement of Trp69 with lysine in either the 56- or 63-kDa gp4 results in defective primer delivery with other functions unaffected. DNA primase harboring lysine at position 69 fails to stabilize the primer on DNA. Thus, a primase subdomain not directly involved in primer synthesis is involved in primer delivery. The stabilization of the primer by DNA primase is necessary for DNA polymerase to initiate synthesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Frick DN, Richardson CC. DNA primases. Annu Rev Biochem. 2001;70:39–80. - PubMed
-
- Hamdan SM, Richardson CC. Motors, switches, and contacts in the replisome. Annu Rev Biochem. 2009;78:205–243. - PubMed
-
- Lee JB, et al. DNA primase acts as a molecular brake in DNA replication. Nature. 2006;439:621–624. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
