

Microtubule organization by the antagonistic mitotic motors kinesin-5 and kinesin-14
- PMID: 20439998
- PMCID: PMC2867311
- DOI: 10.1083/jcb.200910125
Microtubule organization by the antagonistic mitotic motors kinesin-5 and kinesin-14
Abstract
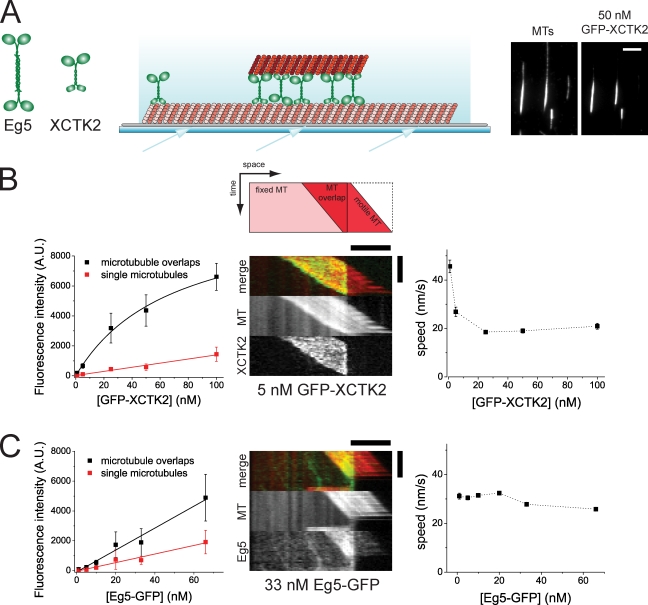
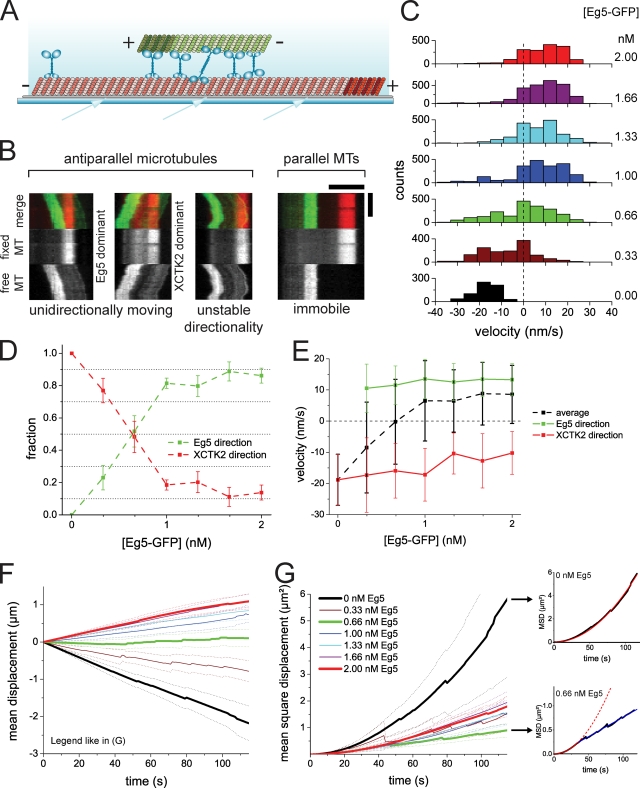
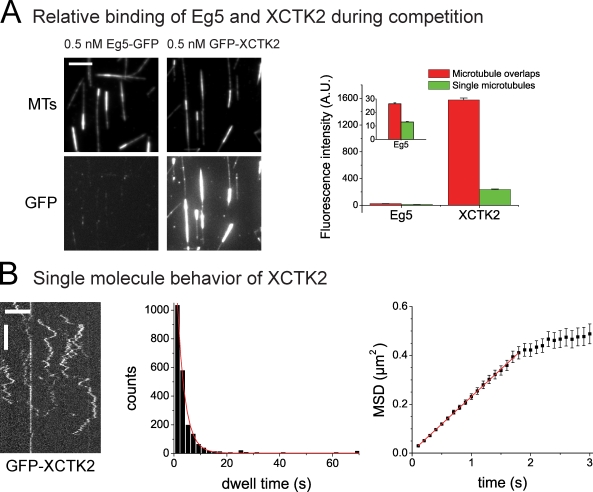
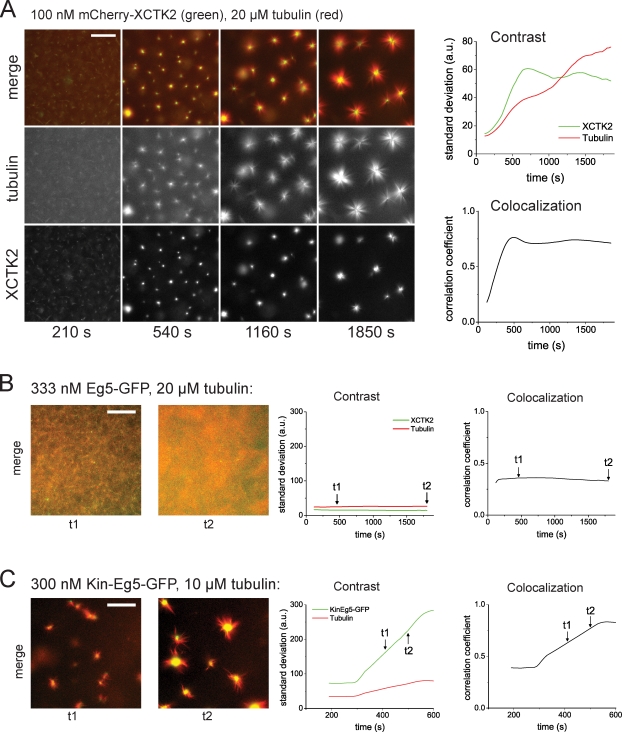
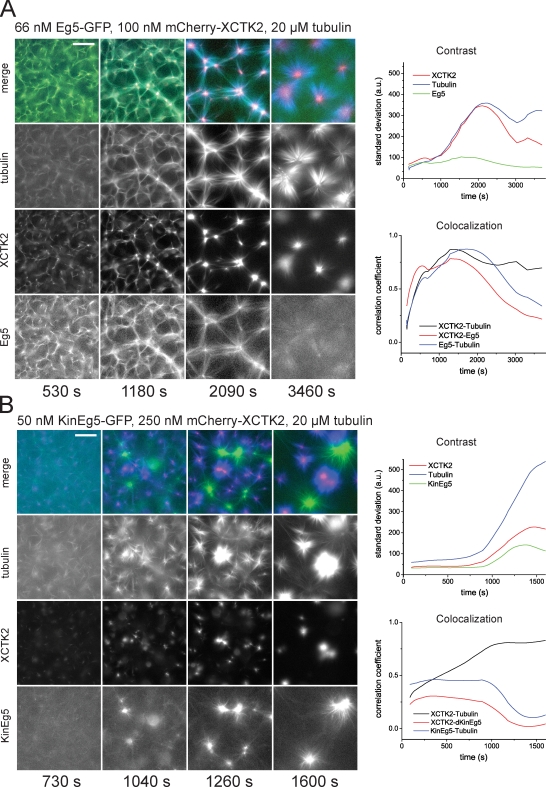
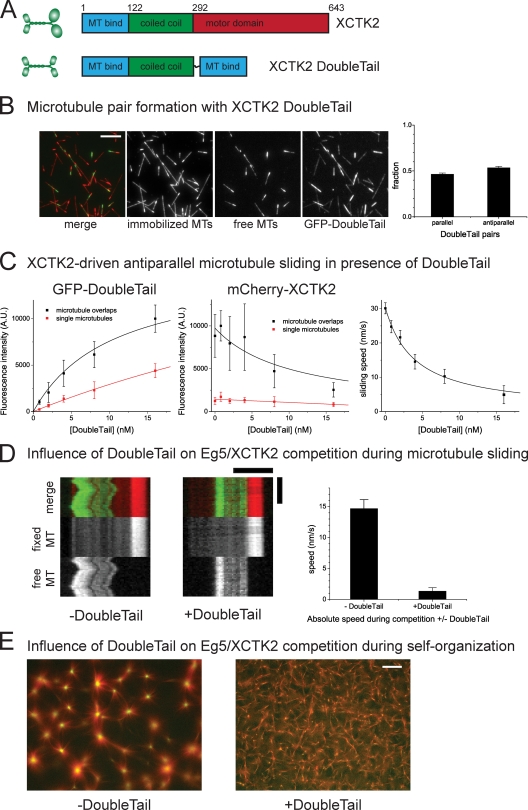
During cell division, different molecular motors act synergistically to rearrange microtubules. Minus end-directed motors are thought to have a dual role: focusing microtubule ends to poles and establishing together with plus end-directed motors a balance of force between antiparallel microtubules in the spindle. We study here the competing action of Xenopus laevis kinesin-14 and -5 in vitro in situations in which these motors with opposite directionality cross-link and slide microtubules. We find that full-length kinesin-14 can form microtubule asters without additional factors, whereas kinesin-5 does not, likely reflecting an adaptation to mitotic function. A stable balance of force is not established between two antiparallel microtubules with these motors. Instead, directional instability is generated, promoting efficient motor and microtubule sorting. A nonmotor microtubule cross-linker can suppress directional instability but also impedes microtubule sorting, illustrating a conflict between stability and dynamicity of organization. These results establish the basic organizational properties of these antagonistic mitotic motors and a microtubule bundler.
Figures
References
-
- Berg H.C. 1993. Random Walks in Biology. Expanded edition Princeton University Press, Princeton, N.J: 152 pp
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
