

A new model for SOS-induced mutagenesis: how RecA protein activates DNA polymerase V
- PMID: 20441441
- PMCID: PMC2874081
- DOI: 10.3109/10409238.2010.480968
A new model for SOS-induced mutagenesis: how RecA protein activates DNA polymerase V
Abstract
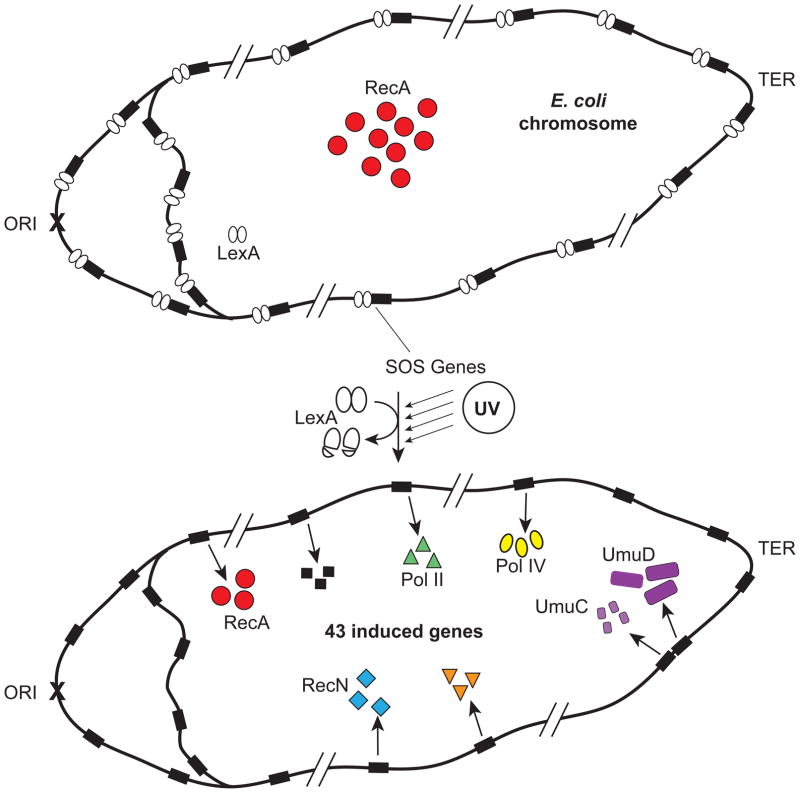
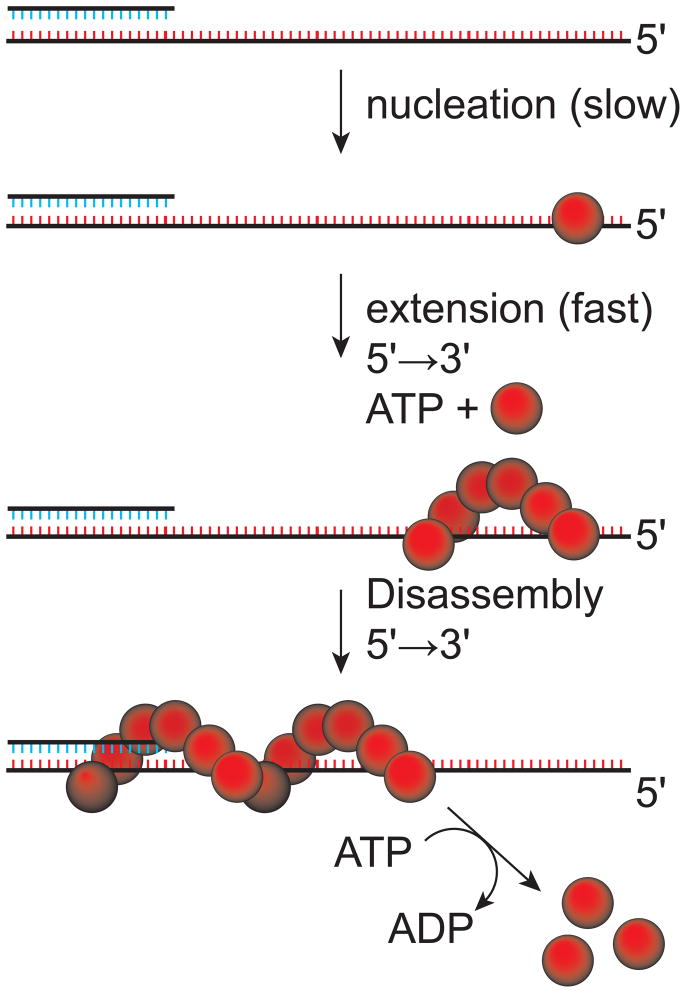
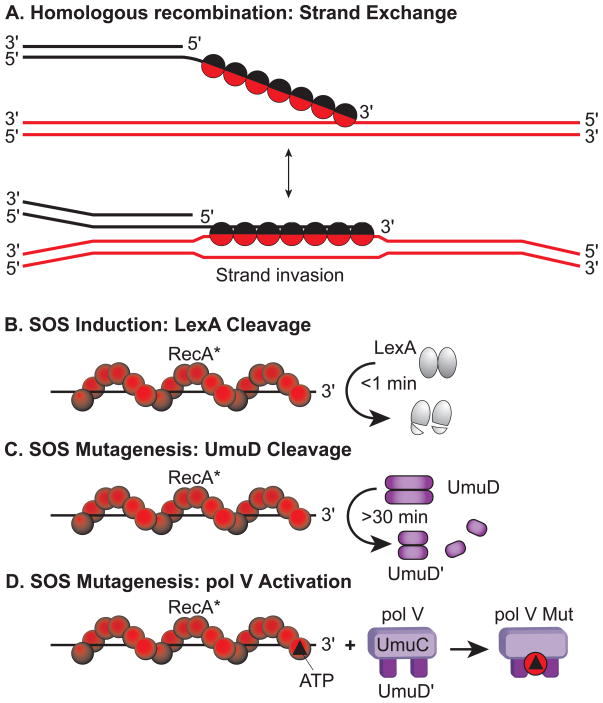
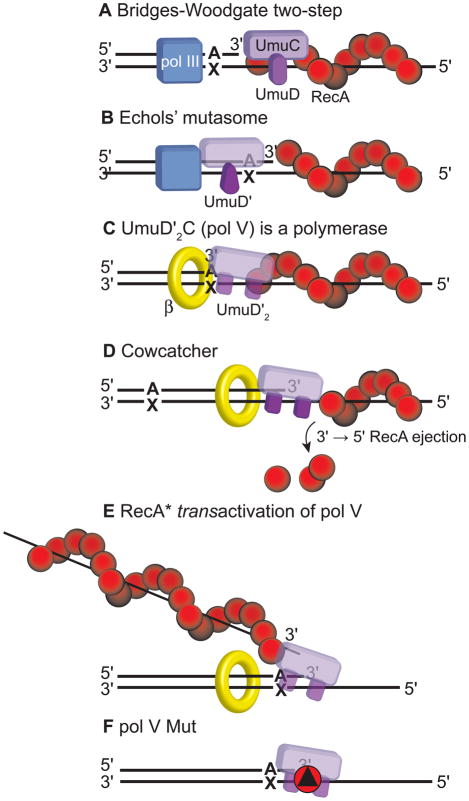
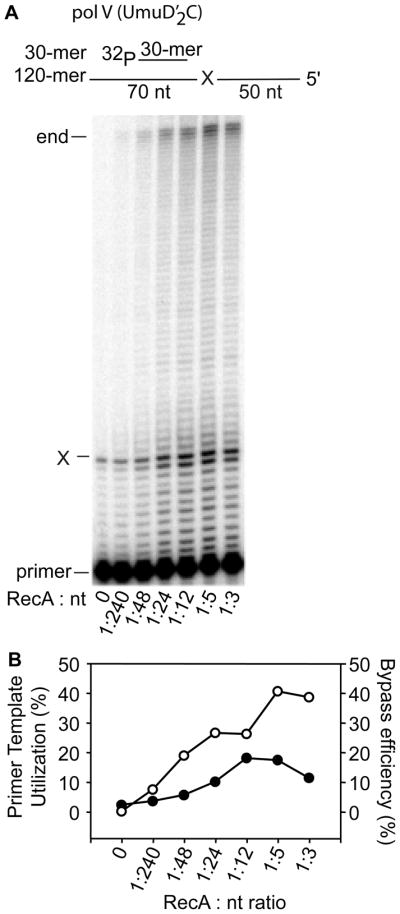
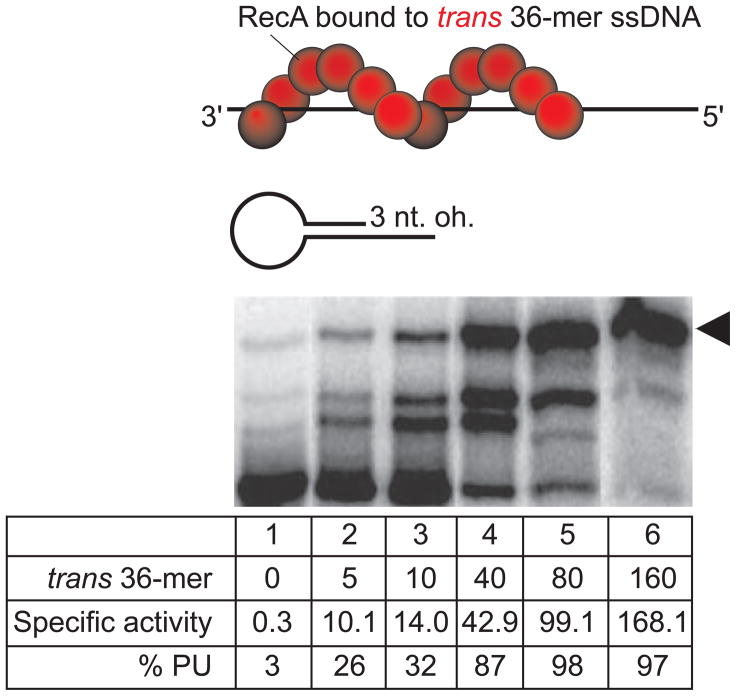
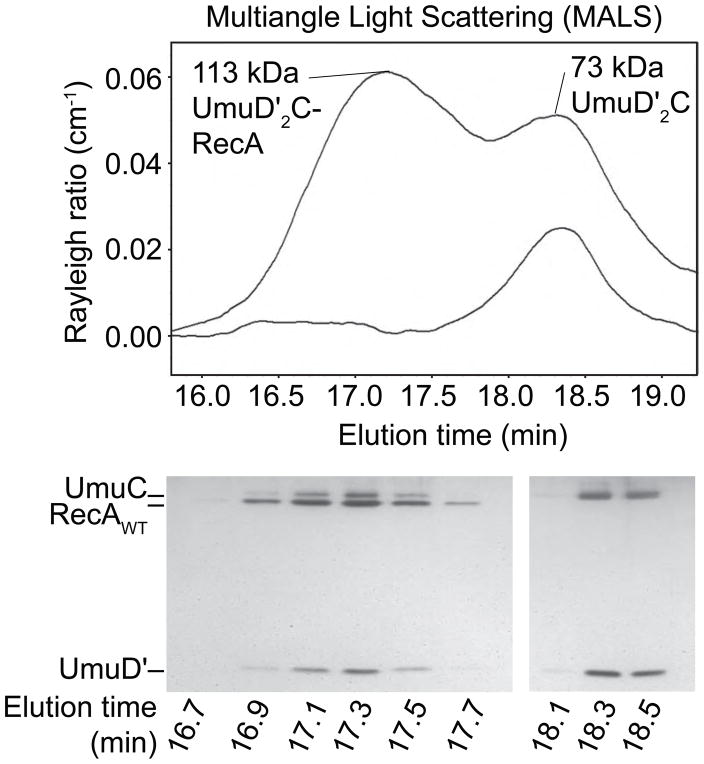
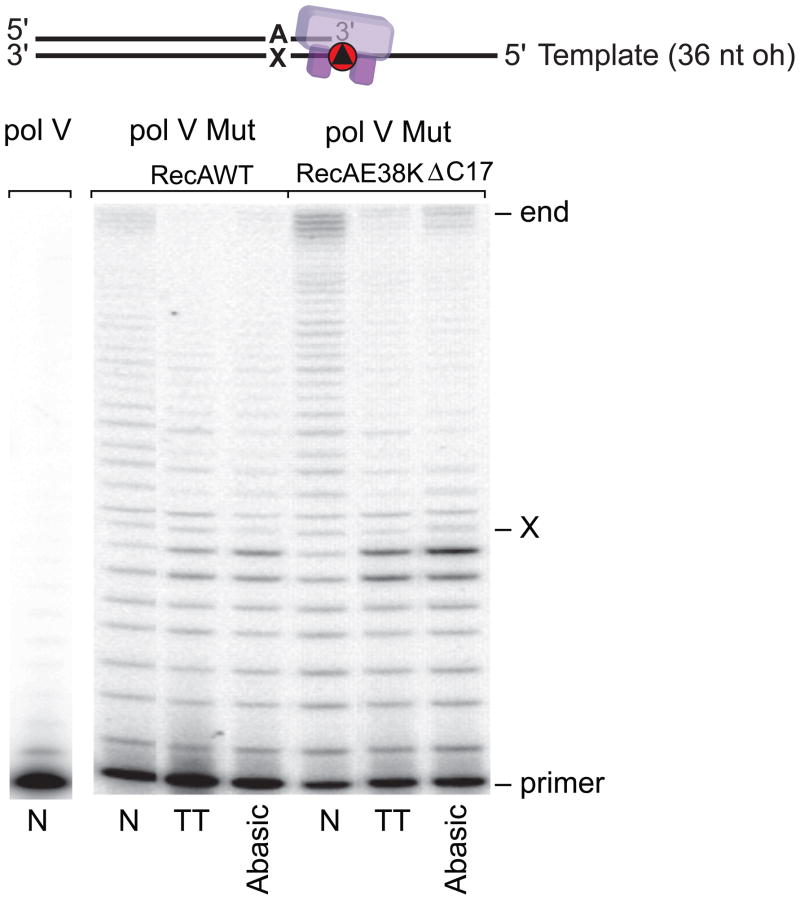
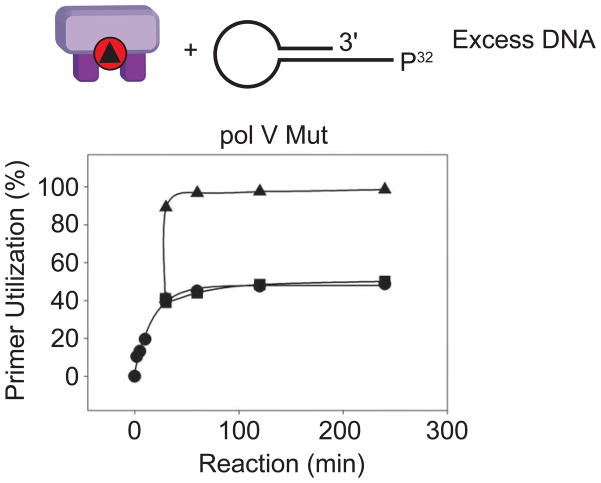
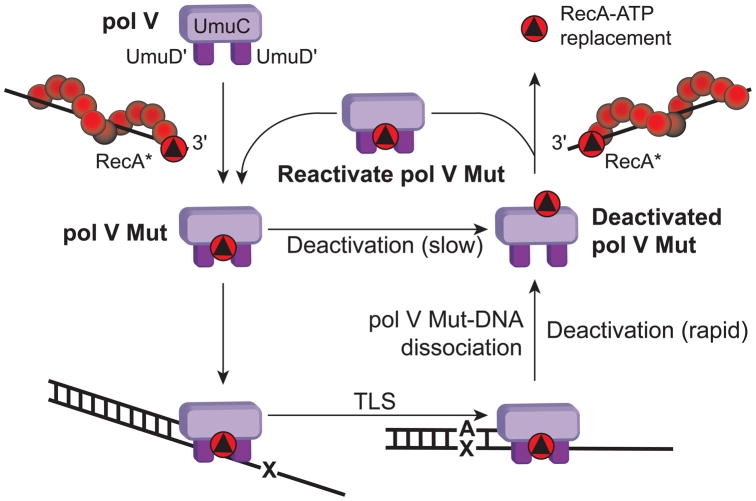
In Escherichia coli, cell survival and genomic stability after UV radiation depends on repair mechanisms induced as part of the SOS response to DNA damage. The early phase of the SOS response is mostly dominated by accurate DNA repair, while the later phase is characterized with elevated mutation levels caused by error-prone DNA replication. SOS mutagenesis is largely the result of the action of DNA polymerase V (pol V), which has the ability to insert nucleotides opposite various DNA lesions in a process termed translesion DNA synthesis (TLS). Pol V is a low-fidelity polymerase that is composed of UmuD'(2)C and is encoded by the umuDC operon. Pol V is strictly regulated in the cell so as to avoid genomic mutation overload. RecA nucleoprotein filaments (RecA*), formed by RecA binding to single-stranded DNA with ATP, are essential for pol V-catalyzed TLS both in vivo and in vitro. This review focuses on recent studies addressing the protein composition of active DNA polymerase V, and the role of RecA protein in activating this enzyme. Based on unforeseen properties of RecA*, we describe a new model for pol V-catalyzed SOS-induced mutagenesis.
Figures
References
-
- Arenson TA, Tsodikov OV, Cox MM. Quantitative analysis of the kinetics of end-dependent disassembly of RecA filaments from ssDNA. J Mol Biol. 1999;288:391–401. - PubMed
-
- Bienko M, Green CM, Crosetto N, Rudolf F, Zapart G, Coull B, Kannouche P, Wider G, Peter M, Lehmann AR, Hofmann K, Dikic I. Ubiquitin-binding domains in Y-family polymerases regulate translesion synthesis. Science. 2005;310:1821–4. - PubMed
-
- Bishop DK, Park D, Xu L, Kleckner N. DMC1: a meiosis-specific yeast homolog of E. coli recA required for recombination, synaptonemal complex formation, and cell cycle progression. Cell. 1992;69:439–56. - PubMed
-
- Bork JM, Cox MM, Inman RB. RecA protein filaments disassemble in the 5′ to 3′ direction on single-stranded DNA. Journal of Biological Chemistry. 2001;276:45740–43. - PubMed
-
- Boudsocq F, Campbell M, Devoret R, Bailone A. Quantitation of the inhibition of Hfr × F- recombination by the mutagenesis complex UmuD′C. J Mol Biol. 1997;270:201–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases