

The kinetics of cooperative cofilin binding reveals two states of the cofilin-actin filament
- PMID: 20441753
- PMCID: PMC2862197
- DOI: 10.1016/j.bpj.2010.01.023
The kinetics of cooperative cofilin binding reveals two states of the cofilin-actin filament
Abstract
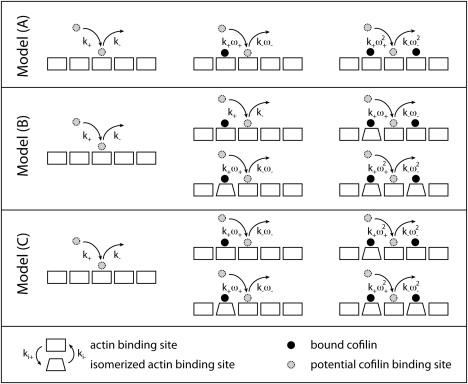
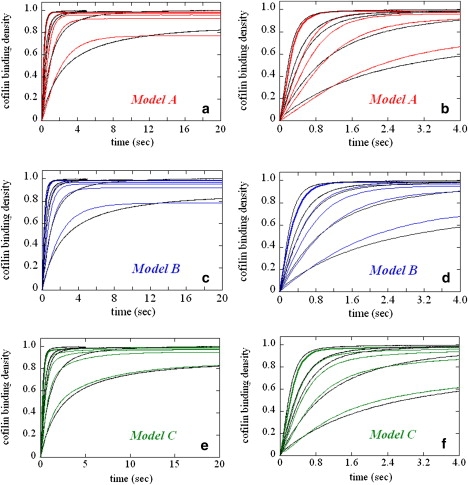
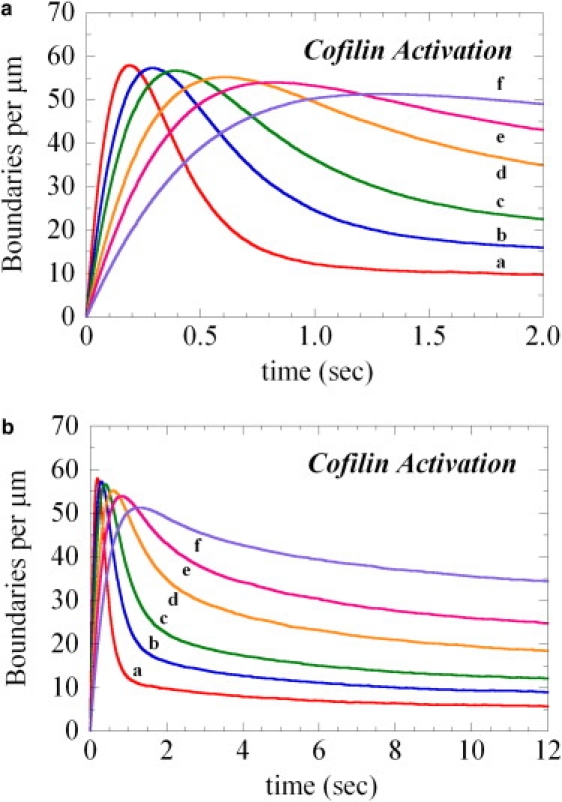
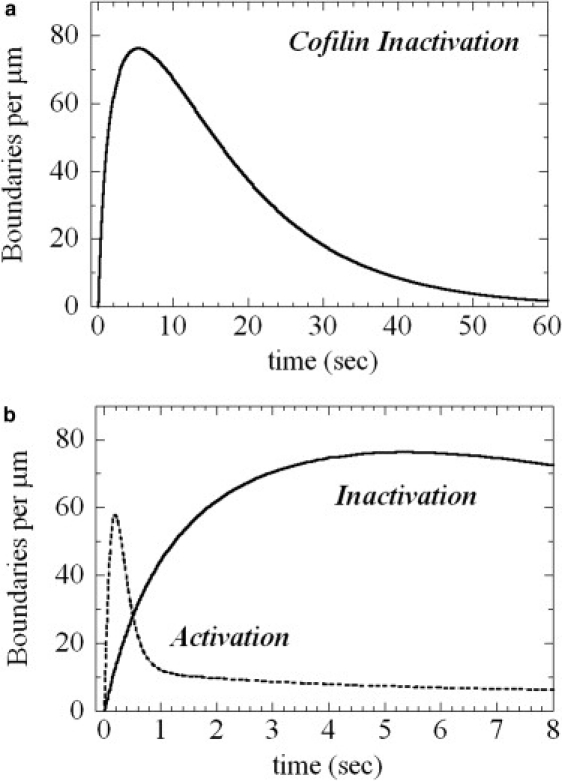
The interaction of cofilin with actin filaments displays positive cooperativity. The equilibrium binding and associated thermodynamic properties of this interaction are well described by a simple, one-dimensional Ising model with nearest neighbor interactions. Here we evaluate the kinetic contributions to cooperative binding and the ability of this model to account for binding across a wide range of cofilin concentrations. A Monte Carlo-based simulation protocol that allows for nearest-neighbor interactions between adjacent binding sites was used to globally fit time courses of human cofilin binding to human nonmuscle (beta-, gamma-) actin filaments. Several extensions of the one-dimensional Ising model were tested, and a mechanism that includes isomerization of the actin filament was found to best account for time courses of association as well as irreversible dissociation from a saturated filament. This model predicts two equilibrium states of the cofilin-actin, or cofilactin, filament, and the resulting set of binding parameters are in agreement with equilibrium thermodynamic parameters. We conclude that despite its simplicity, this one-dimensional Ising model is a reliable model for analyzing and interpreting the energetics and kinetics of cooperative cofilin-actin filament interactions. The model predicts that severing activity associated with boundaries between bare and decorated segments will not be linear, but display a transient burst at short times on cofilin activation then dissipate due to a kinetic competition between severing activity and cofilin binding. A second peak of severing activity is predicted to arise from irreversible cofilin dissociation on inactivation. These behaviors predict what we believe to be novel mechanisms of cofilin severing and spatial regulation of actin filament turnover in cells. The methods developed for this system are generally applicable to the kinetic analysis of cooperative ligand binding to linear polymers.
Copyright (c) 2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures



Similar articles
-
Structures of cofilin-induced structural changes reveal local and asymmetric perturbations of actin filaments.Proc Natl Acad Sci U S A. 2020 Jan 21;117(3):1478-1484. doi: 10.1073/pnas.1915987117. Epub 2020 Jan 3. Proc Natl Acad Sci U S A. 2020. PMID: 31900364 Free PMC article.
-
Actin Filament Strain Promotes Severing and Cofilin Dissociation.Biophys J. 2017 Jun 20;112(12):2624-2633. doi: 10.1016/j.bpj.2017.05.016. Biophys J. 2017. PMID: 28636918 Free PMC article.
-
Molecular origins of cofilin-linked changes in actin filament mechanics.J Mol Biol. 2013 Apr 12;425(7):1225-40. doi: 10.1016/j.jmb.2013.01.020. Epub 2013 Jan 24. J Mol Biol. 2013. PMID: 23352932 Free PMC article.
-
Biophysics of actin filament severing by cofilin.FEBS Lett. 2013 Apr 17;587(8):1215-9. doi: 10.1016/j.febslet.2013.01.062. Epub 2013 Feb 5. FEBS Lett. 2013. PMID: 23395798 Free PMC article. Review.
-
Tropomyosin and ADF/cofilin as collaborators and competitors.Adv Exp Med Biol. 2008;644:232-49. doi: 10.1007/978-0-387-85766-4_18. Adv Exp Med Biol. 2008. PMID: 19209826 Review.
Cited by
-
Structures of cofilin-induced structural changes reveal local and asymmetric perturbations of actin filaments.Proc Natl Acad Sci U S A. 2020 Jan 21;117(3):1478-1484. doi: 10.1073/pnas.1915987117. Epub 2020 Jan 3. Proc Natl Acad Sci U S A. 2020. PMID: 31900364 Free PMC article.
-
Actin filament remodeling by actin depolymerization factor/cofilin.Proc Natl Acad Sci U S A. 2010 Apr 20;107(16):7299-304. doi: 10.1073/pnas.0911675107. Epub 2010 Apr 5. Proc Natl Acad Sci U S A. 2010. PMID: 20368459 Free PMC article.
-
Cofilin-induced unidirectional cooperative conformational changes in actin filaments revealed by high-speed atomic force microscopy.Elife. 2015 Feb 2;4:e04806. doi: 10.7554/eLife.04806. Elife. 2015. PMID: 25642645 Free PMC article.
-
Srv2/cyclase-associated protein forms hexameric shurikens that directly catalyze actin filament severing by cofilin.Mol Biol Cell. 2013 Jan;24(1):31-41. doi: 10.1091/mbc.E12-08-0589. Epub 2012 Nov 7. Mol Biol Cell. 2013. PMID: 23135996 Free PMC article.
-
Building a dendritic actin filament network branch by branch: models of filament orientation pattern and force generation in lamellipodia.Biophys Rev. 2018 Dec;10(6):1577-1585. doi: 10.1007/s12551-018-0475-7. Epub 2018 Nov 12. Biophys Rev. 2018. PMID: 30421277 Free PMC article. Review.
References
-
- Michelot A., Berro J., Blanchoin L. Actin-filament stochastic dynamics mediated by ADF/cofilin. Curr. Biol. 2007;17:825–833. - PubMed
-
- Andrianantoandro E., Pollard T.D. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Mol. Cell. 2006;24:13–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
