

NMR solution structure of poliovirus uridylyated peptide linked to the genome (VPgpU)
- PMID: 20441784
- PMCID: PMC2905501
- DOI: 10.1016/j.peptides.2010.04.021
NMR solution structure of poliovirus uridylyated peptide linked to the genome (VPgpU)
Abstract
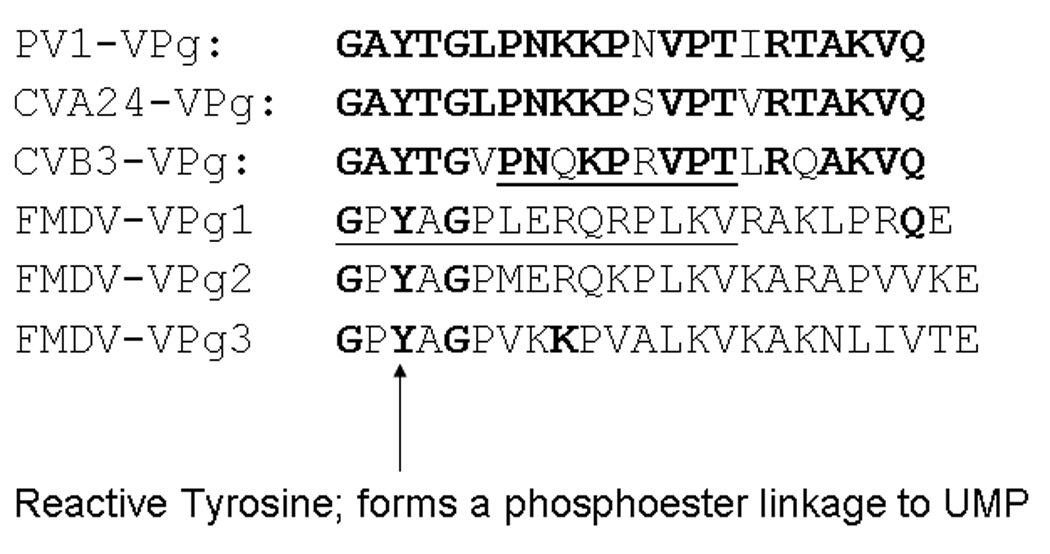
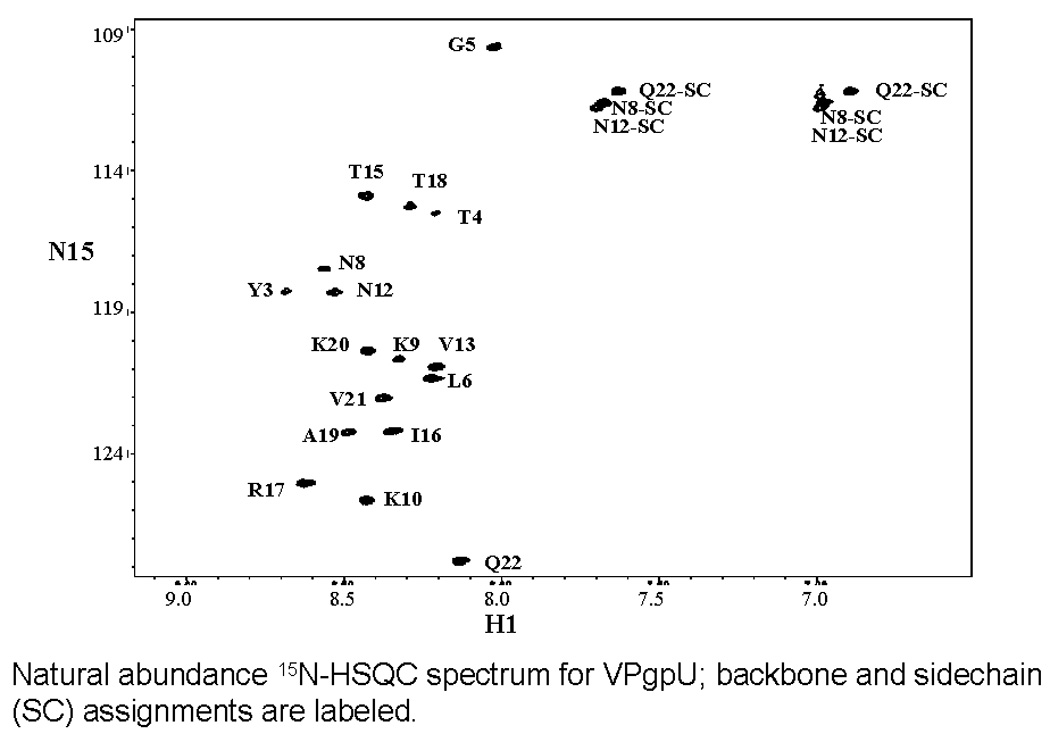
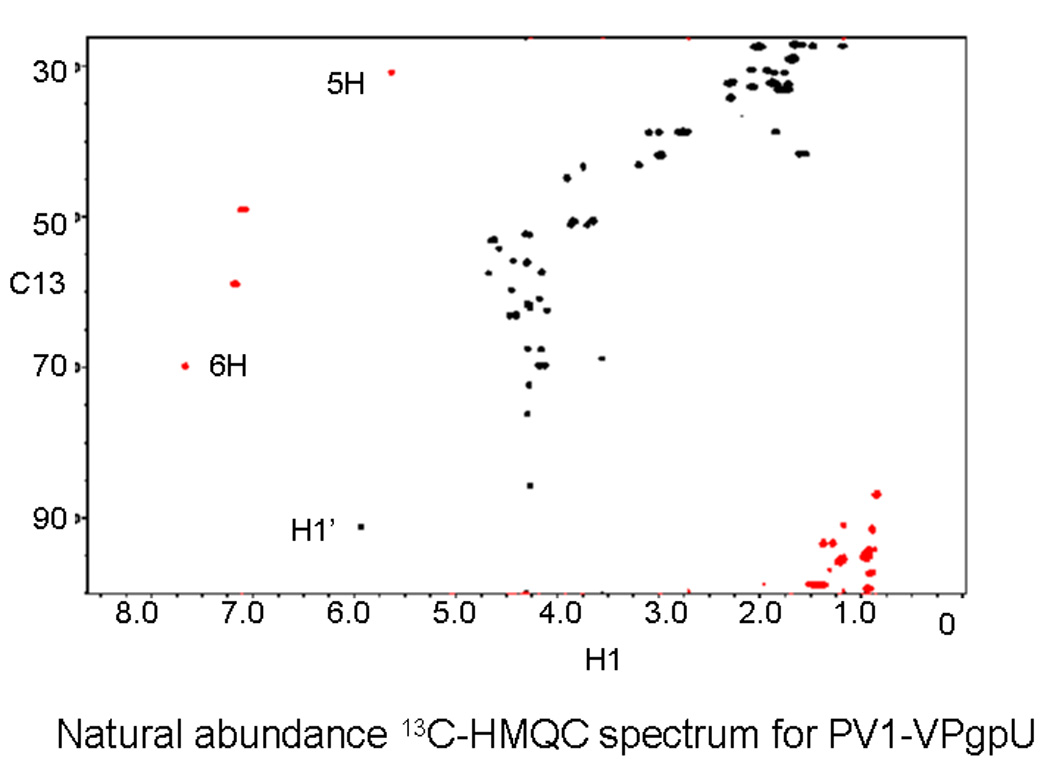
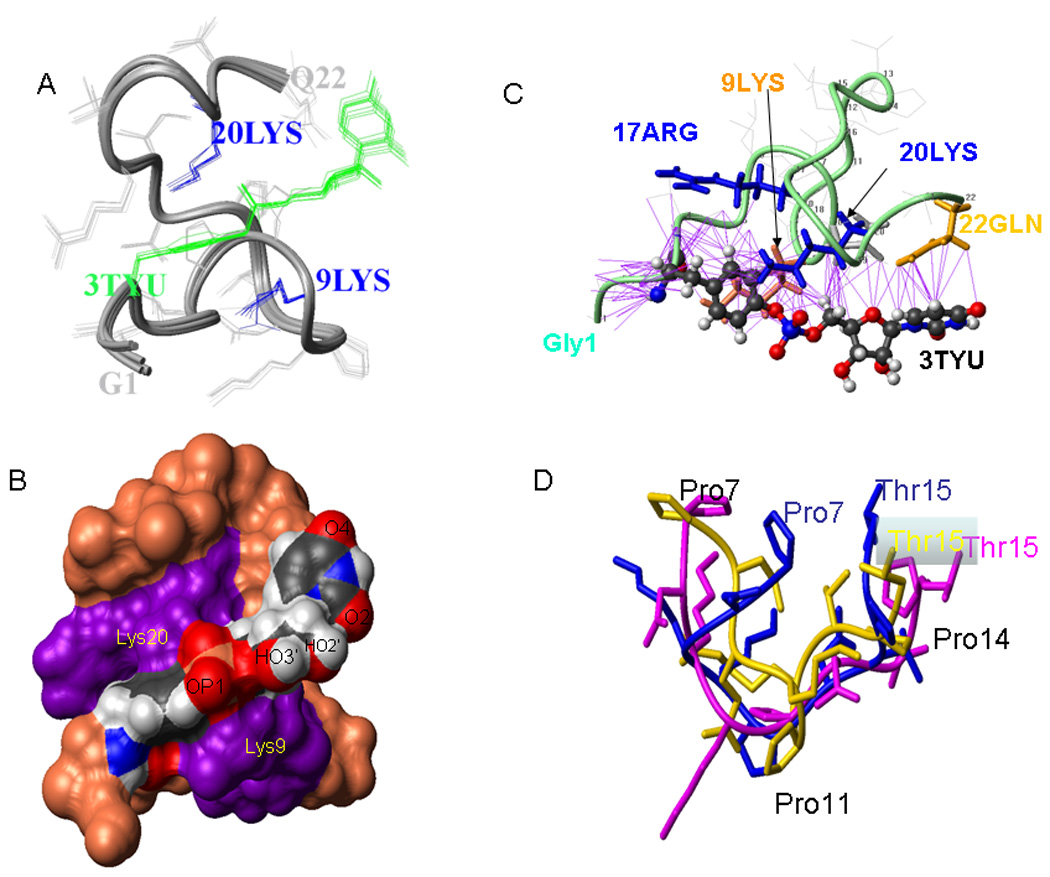
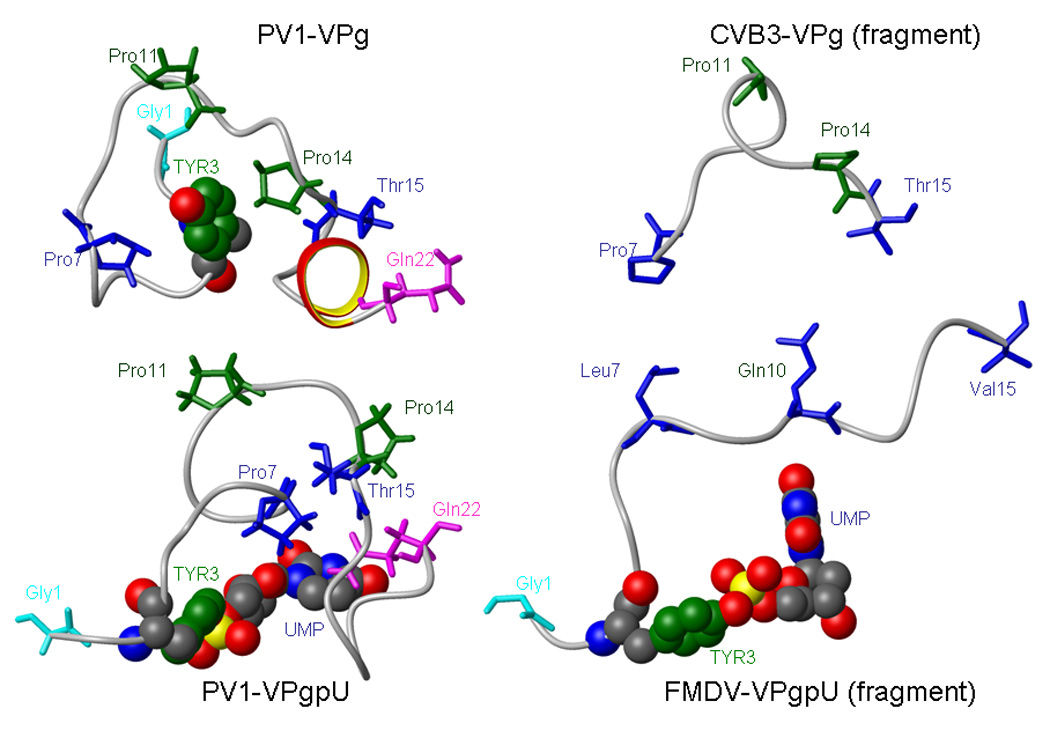
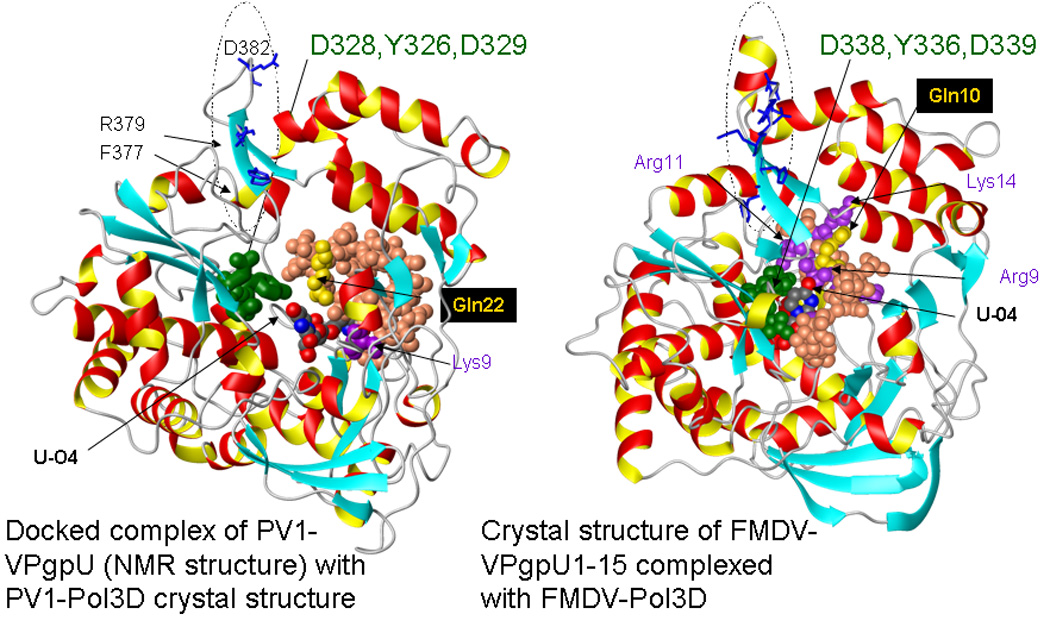
Picornaviruses have a 22-24 amino acid peptide, VPg, bound covalently at the 5' end of their RNA, that is essential for replication. VPgs are uridylylated at a conserved tyrosine to form VPgpU, the primer of RNA synthesis by the viral polymerase. This first complete structure for any uridylylated VPg, of poliovirus type 1 (PV1)-VPgpU, shows that conserved amino acids in VPg stabilize the bound UMP, with the uridine atoms involved in base pairing and chain elongation projected outward. Comparing this structure to PV1-VPg and partial structures of VPg/VPgpU from other picornaviruses suggests that enteroviral polymerases require a more stable VPg structure than does the distantly related aphthovirus, foot and mouth disease virus (FMDV). The glutamine residue at the C-terminus of PV1-VPgpU lies in back of the uridine base and may stabilize its position during chain elongation and/or contribute to base specificity. Under in vivo-like conditions with the authentic cre(2C) hairpin RNA and Mg(2+), 5-methylUTP cannot compete with UTP for VPg uridylyation in an in vitro uridylyation assay, but both nucleotides are equally incorporated by PV1-polymerase with Mn(2+) and a poly-A RNA template. This indicates the 5 position is recognized under in vivo conditions. The compact VPgpU structure docks within the active site cavity of the PV-polymerase, close to the position seen for the fragment of FMDV-VPgpU with its polymerase. This structure could aid in design of novel enterovirus inhibitors, and stabilization upon uridylylation may also be pertinent for post-translational uridylylation reactions that underlie other biological processes.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures


Similar articles
-
Sequences and Structures of Viral Proteins Linked to the Genomes (VPg) of RNA Viruses.Viruses. 2025 Apr 29;17(5):645. doi: 10.3390/v17050645. Viruses. 2025. PMID: 40431656 Free PMC article. Review.
-
(F)uridylylated Peptides Linked to VPg1 of Foot-and- Mouth Disease Virus (FMDV): Design, Synthesis and X-Ray Crystallography of the Complexes with FMDV RNA-Dependent RNA Polymerase.Molecules. 2019 Jun 26;24(13):2360. doi: 10.3390/molecules24132360. Molecules. 2019. PMID: 31247979 Free PMC article.
-
Identification of an RNA hairpin in poliovirus RNA that serves as the primary template in the in vitro uridylylation of VPg.J Virol. 2000 Nov;74(22):10359-70. doi: 10.1128/jvi.74.22.10359-10370.2000. J Virol. 2000. PMID: 11044080 Free PMC article.
-
Genetic and biochemical studies of poliovirus cis-acting replication element cre in relation to VPg uridylylation.J Virol. 2000 Nov;74(22):10371-80. doi: 10.1128/jvi.74.22.10371-10380.2000. J Virol. 2000. PMID: 11044081 Free PMC article.
-
Formation and working mechanism of the picornavirus VPg uridylylation complex.Curr Opin Virol. 2014 Dec;9:24-30. doi: 10.1016/j.coviro.2014.09.003. Epub 2014 Sep 19. Curr Opin Virol. 2014. PMID: 25240314 Review.
Cited by
-
Allosteric inhibitors of Coxsackie virus A24 RNA polymerase.Bioorg Med Chem. 2016 Feb 15;24(4):570-7. doi: 10.1016/j.bmc.2015.12.023. Epub 2015 Dec 15. Bioorg Med Chem. 2016. PMID: 26762834 Free PMC article.
-
The twenty-nine amino acid C-terminal cytoplasmic domain of poliovirus 3AB is critical for nucleic acid chaperone activity.RNA Biol. 2010 Nov-Dec;7(6):820-9. doi: 10.4161/rna.7.6.13781. Epub 2010 Nov 1. RNA Biol. 2010. PMID: 21045553 Free PMC article.
-
Picornaviral polymerase structure, function, and fidelity modulation.Virus Res. 2017 Apr 15;234:4-20. doi: 10.1016/j.virusres.2017.01.026. Epub 2017 Feb 2. Virus Res. 2017. PMID: 28163093 Free PMC article. Review.
-
Enterovirus infections of the central nervous system.Virology. 2011 Mar 15;411(2):288-305. doi: 10.1016/j.virol.2010.12.014. Epub 2011 Jan 20. Virology. 2011. PMID: 21251690 Free PMC article. Review.
-
Sequences and Structures of Viral Proteins Linked to the Genomes (VPg) of RNA Viruses.Viruses. 2025 Apr 29;17(5):645. doi: 10.3390/v17050645. Viruses. 2025. PMID: 40431656 Free PMC article. Review.
References
-
- Andino R, Rieckhof GE, Baltimore D. A Functional Ribonucleoprotein Complex Forms around the 5' End of Poliovirus Rna. Cell. 1990;63:369–380. - PubMed
-
- Arnold JJ, Gosh KB, Cameron C. Poliovirus RNA-dependent RNA polymerase 3Dpol. Divalent cation modulation of primer, template and nucleotide selection. J. Biol. Chem. 1999;274:37060–37069. - PubMed
-
- Barreteau H, Kova A, Boniface DA, Sova M, Gobec S, Blanot D. Cytoplasmic steps of peptidoglycan biosynthesis. FEMS Microbiology Reviews. 2008;32:168–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
