

Membrane wounding triggers ATP release and dysferlin-mediated intercellular calcium signaling
- PMID: 20442251
- PMCID: PMC2873225
- DOI: 10.1242/jcs.066084
Membrane wounding triggers ATP release and dysferlin-mediated intercellular calcium signaling
Abstract
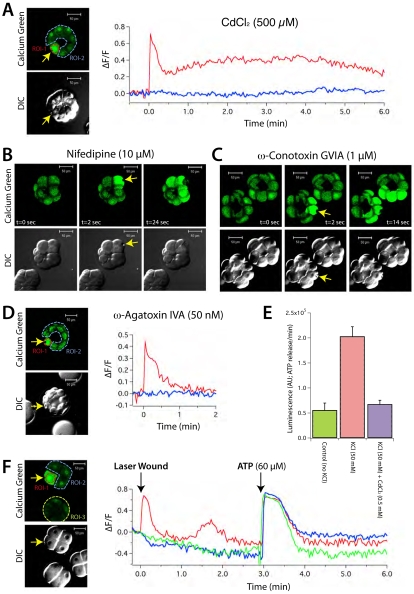
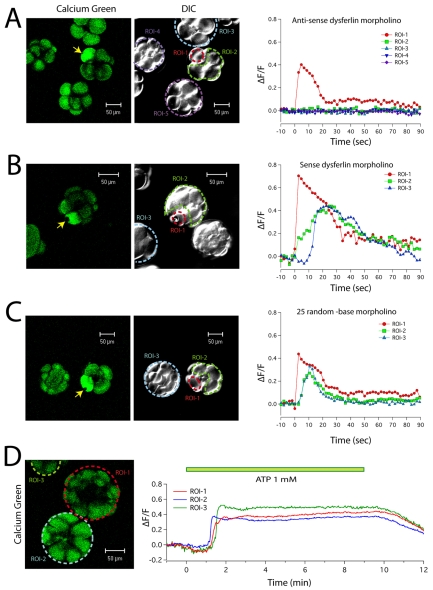
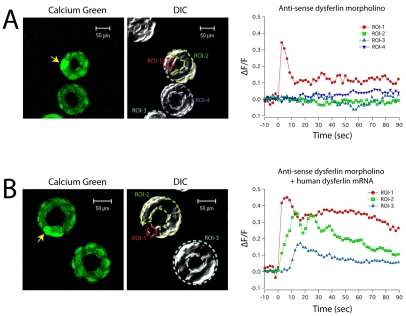
Dysferlin is a Ca(2+)-binding protein found in many different cell types. It is required for membrane wound repair in muscle, but it is not known whether it has the same function in other cells. Here we report the activation of an intercellular signaling pathway in sea urchin embryos by membrane wounding that evokes Ca(2+) spikes in neighboring cells. This pathway was mimicked by ATP application, and inhibited by apyrase, cadmium, and omega-agatoxin-IVA. Microinjection of dysferlin antisense phosphorodiamidate morpholino oligonucleotides blocked this pathway, whereas control morpholinos did not. Co-injection of mRNA encoding human dysferlin with the inhibitory morpholino rescued signaling activity. We conclude that in sea urchin embryos dysferlin mediates Ca(2+)-triggered intercellular signaling in response to membrane wounding.
Figures




References
-
- Angerer L. M., Angerer R. C. (2004). Disruption of gene function using antisense morpholinos. In Development of Sea Urchins, Ascidians, and other Invertebrate Deuterosomes: Experimental Approaches, vol. 74 (ed. Ettensohn C. A., Wessel G. M., Wray G. A.), p. 883 Amsterdam: Elsevier;
-
- Anselmi F., Hernandez V. H., Crispino G., Seydel A., Ortolano S., Roper S. D., Kessaris N., Richardson W., Rickheit G., Filippov M. A., et al. (2008). ATP release through connexin hemichannels and gap junction transfer of second messengers propagate Ca2+ signals across the inner ear. Proc. Natl. Acad. Sci. USA 105, 18770-18775 - PMC - PubMed
-
- Aoki M., Liu J., Richard I., Bashir R., Britton S., Keers S. M., Oeltjen J., Brown H. E., Marchand S., Bourg N., et al. (2001). Genomic organization of the dysferlin gene and novel mutations in Miyoshi myopathy. Neurology 57, 271-278 - PubMed
-
- Bansal D., Miyake K., Vogel S. S., Groh S., Chen C. C., Williamson R., McNeil P. L., Campbell K. P. (2003). Defective membrane repair in dysferlin-deficient muscular dystrophy. Nature 423, 168-172 - PubMed
-
- Bashir R., Strachan T., Keers S., Stephenson A., Mahjneh I., Marconi G., Nashef L., Bushby K. M. (1994). A gene for autosomal recessive limb-girdle muscular dystrophy maps to chromosome 2p. Hum. Mol. Genet. 3, 455-457 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
