

Dynamic link of DNA demethylation, DNA strand breaks and repair in mouse zygotes
- PMID: 20442707
- PMCID: PMC2885932
- DOI: 10.1038/emboj.2010.80
Dynamic link of DNA demethylation, DNA strand breaks and repair in mouse zygotes
Abstract
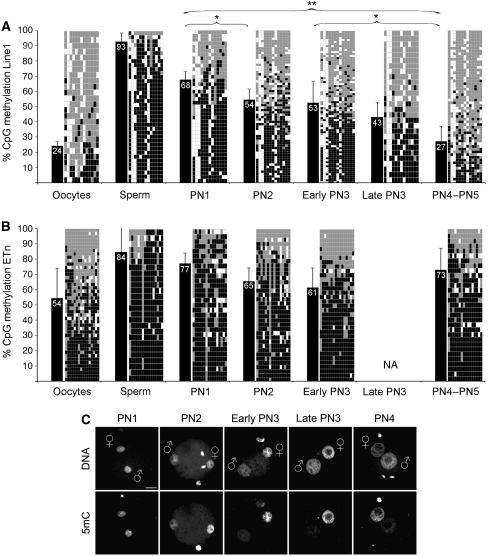
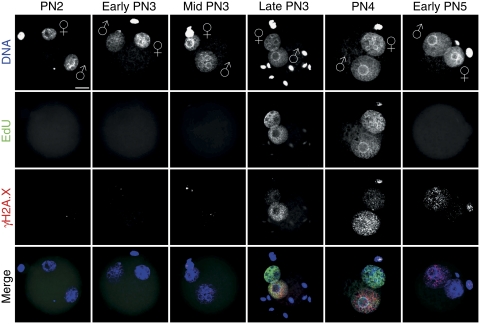
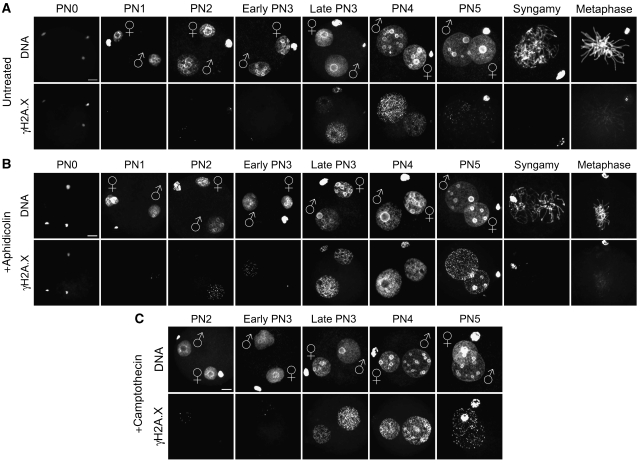
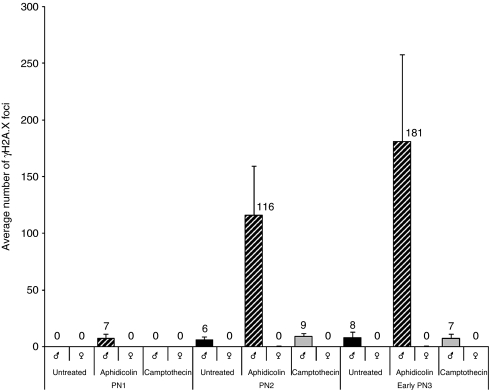
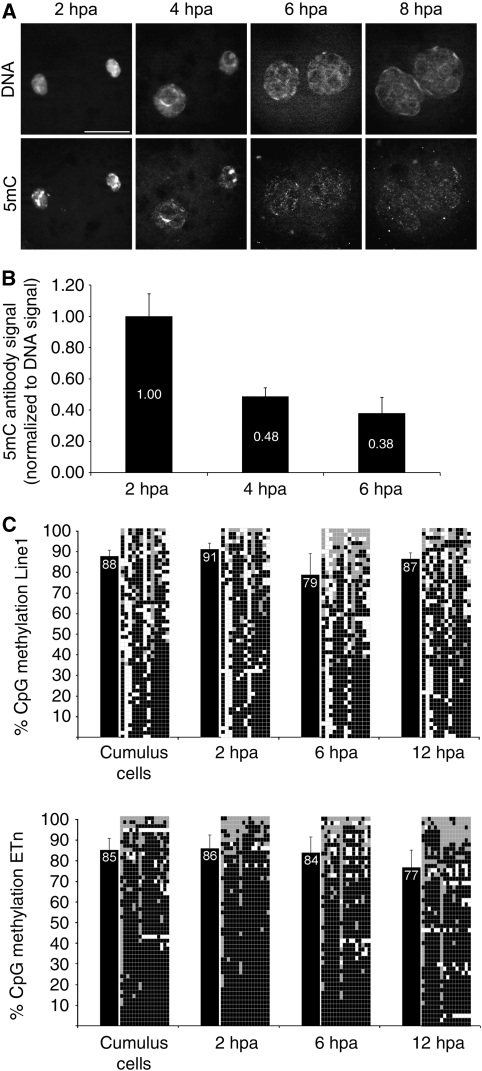
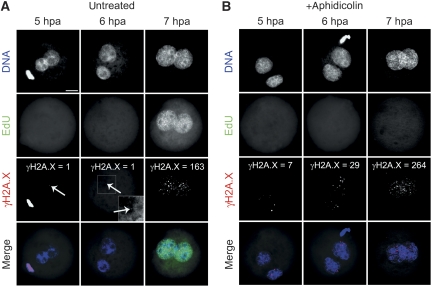
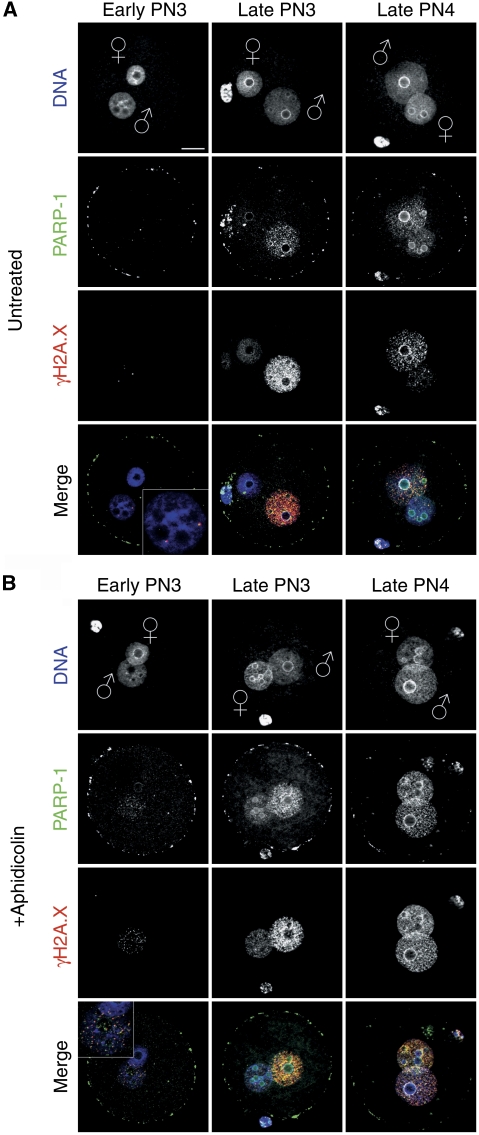
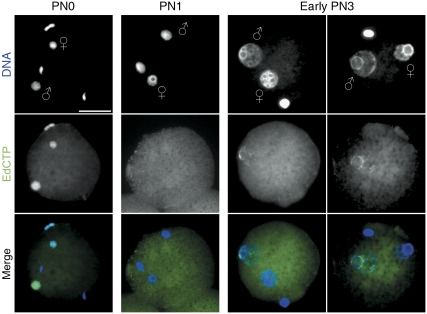
In mammalian zygotes, the 5-methyl-cytosine (5mC) content of paternal chromosomes is rapidly changed by a yet unknown but presumably active enzymatic mechanism. Here, we describe the developmental dynamics and parental asymmetries of DNA methylation in relation to the presence of DNA strand breaks, DNA repair markers and a precise timing of zygotic DNA replication. The analysis shows that distinct pre-replicative (active) and replicative (active and passive) phases of DNA demethylation can be observed. These phases of DNA demethylation are concomitant with the appearance of DNA strand breaks and DNA repair markers such as gammaH2A.X and PARP-1, respectively. The same correlations are found in cloned embryos obtained after somatic cell nuclear transfer. Together, the data suggest that (1) DNA-methylation reprogramming is more complex and extended as anticipated earlier and (2) the DNA demethylation, particularly the rapid loss of 5mC in paternal DNA, is likely to be linked to DNA repair mechanisms.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Adenot PG, Mercier Y, Renard JP, Thompson EM (1997) Differential H4 acetylation of paternal and maternal chromatin precedes DNA replication and differential transcriptional activity in pronuclei of 1-cell mouse embryos. Development 124: 4615–4625 - PubMed
-
- Aoki E, Schultz RM (1999) DNA replication in the 1-cell mouse embryo: stimulatory effect of histone acetylation. Zygote 7: 165–172 - PubMed
-
- Aoki F, Worrad DM, Schultz RM (1997) Regulation of transcriptional activity during the first and second cell cycles in the preimplantation mouse embryo. Dev Biol 181: 296–307 - PubMed
-
- Barreto G, Schafer A, Marhold J, Stach D, Swaminathan SK, Handa V, Doderlein G, Maltry N, Wu W, Lyko F, Niehrs C (2007) Gadd45a promotes epigenetic gene activation by repair-mediated DNA demethylation. Nature 445: 671–675 - PubMed
-
- Berger NA, Kurohara KK, Petzold SJ, Sikorski GW (1979) Aphidicolin inhibits eukaryotic DNA replication and repair—implications for involvement of DNA polymerase alpha in both processes. Biochem Biophys Res Commun 89: 218–225 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
