

Chaperonin containing T-complex polypeptide subunit eta (CCT-eta) is a specific regulator of fibroblast motility and contractility
- PMID: 20442790
- PMCID: PMC2862014
- DOI: 10.1371/journal.pone.0010063
Chaperonin containing T-complex polypeptide subunit eta (CCT-eta) is a specific regulator of fibroblast motility and contractility
Abstract
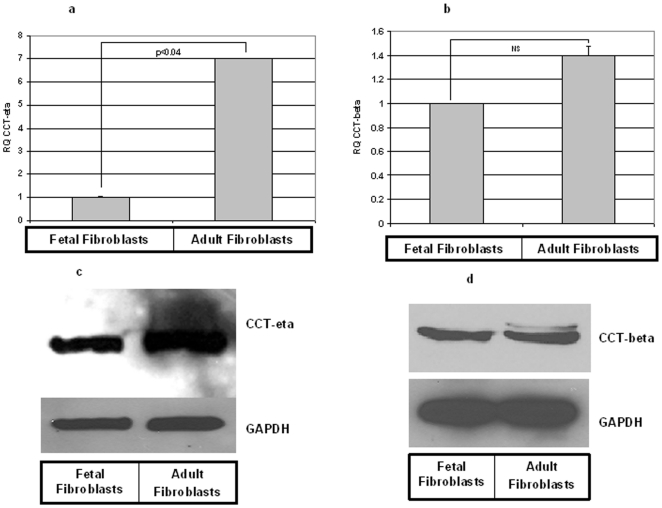
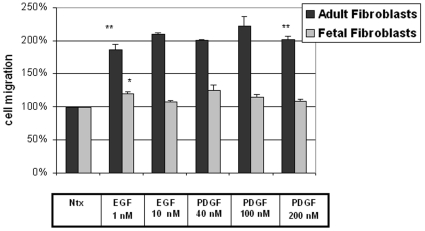
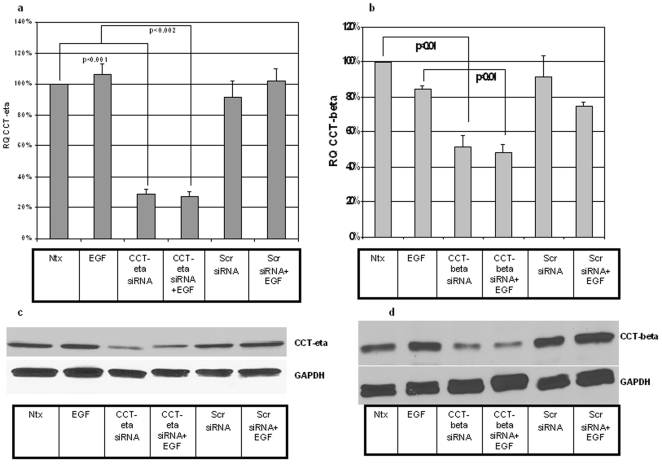
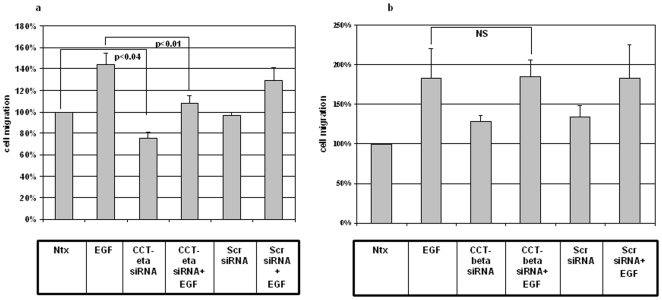
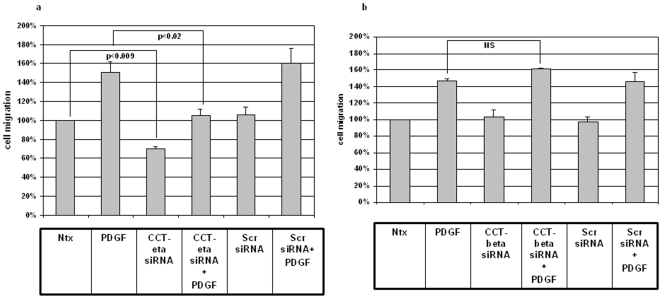
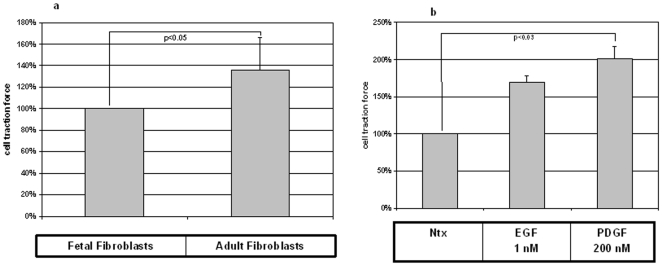
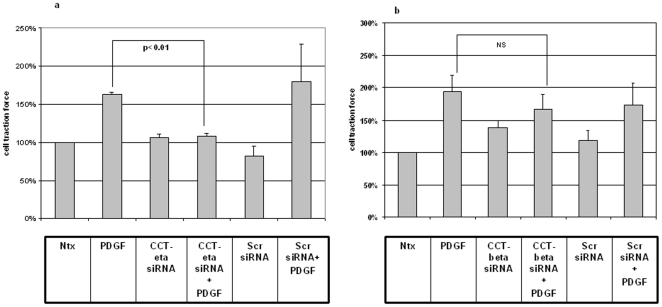
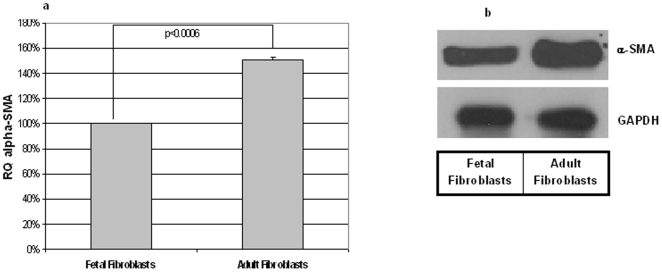
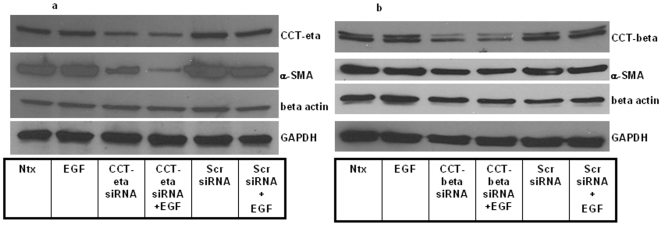
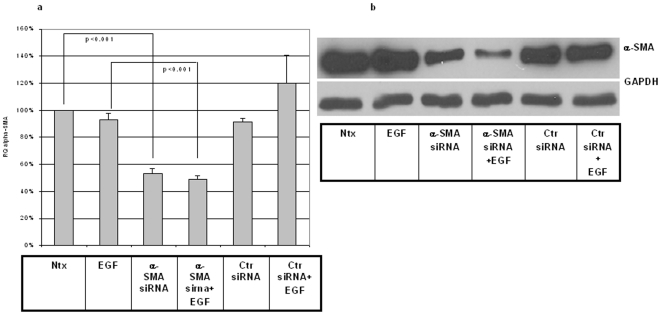
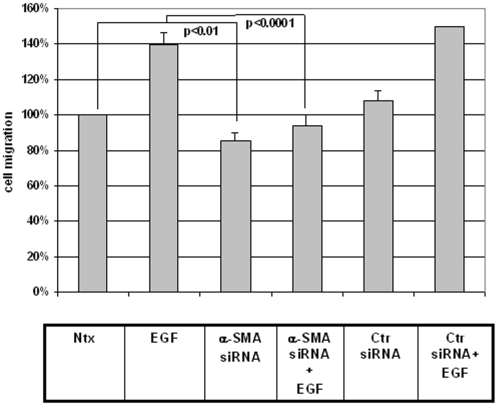
Integumentary wounds in mammalian fetuses heal without scar; this scarless wound healing is intrinsic to fetal tissues and is notable for absence of the contraction seen in postnatal (adult) wounds. The precise molecular signals determining the scarless phenotype remain unclear. We have previously reported that the eta subunit of the chaperonin containing T-complex polypeptide (CCT-eta) is specifically reduced in healing fetal wounds in a rabbit model. In this study, we examine the role of CCT-eta in fibroblast motility and contractility, properties essential to wound healing and scar formation. We demonstrate that CCT-eta (but not CCT-beta) is underexpressed in fetal fibroblasts compared to adult fibroblasts. An in vitro wound healing assay demonstrated that adult fibroblasts showed increased cell migration in response to epidermal growth factor (EGF) and platelet derived growth factor (PDGF) stimulation, whereas fetal fibroblasts were unresponsive. Downregulation of CCT-eta in adult fibroblasts with short inhibitory RNA (siRNA) reduced cellular motility, both basal and growth factor-induced; in contrast, siRNA against CCT-beta had no such effect. Adult fibroblasts were more inherently contractile than fetal fibroblasts by cellular traction force microscopy; this contractility was increased by treatment with EGF and PDGF. CCT-eta siRNA inhibited the PDGF-induction of adult fibroblast contractility, whereas CCT-beta siRNA had no such effect. In each of these instances, the effect of downregulating CCT-eta was to modulate the behavior of adult fibroblasts so as to more closely approximate the characteristics of fetal fibroblasts. We next examined the effect of CCT-eta modulation on alpha-smooth muscle actin (alpha-SMA) expression, a gene product well known to play a critical role in adult wound healing. Fetal fibroblasts were found to constitutively express less alpha-SMA than adult cells. Reduction of CCT-eta with siRNA had minimal effect on cellular beta-actin but markedly decreased alpha-SMA; in contrast, reduction of CCT-beta had minimal effect on either actin isoform. Direct inhibition of alpha-SMA with siRNA reduced both basal and growth factor-induced fibroblast motility. These results indicate that CCT-eta is a specific regulator of fibroblast motility and contractility and may be a key determinant of the scarless wound healing phenotype by means of its specific regulation of alpha-SMA expression.
Conflict of interest statement
Figures
Similar articles
-
Increased CCT-eta expression is a marker of latent and active disease and a modulator of fibroblast contractility in Dupuytren's contracture.Cell Stress Chaperones. 2013 Jul;18(4):397-404. doi: 10.1007/s12192-012-0392-9. Epub 2013 Jan 6. Cell Stress Chaperones. 2013. PMID: 23292503 Free PMC article.
-
RNA differential display of scarless wound healing in fetal rabbit indicates downregulation of a CCT chaperonin subunit and upregulation of a glycophorin-like gene transcript.J Pediatr Surg. 2000 Mar;35(3):406-19. doi: 10.1016/s0022-3468(00)90204-5. J Pediatr Surg. 2000. PMID: 10726679
-
Cloning and expression of rabbit CCT subunits eta and beta in healing cutaneous wounds.Cell Stress Chaperones. 2010 Nov;15(6):819-26. doi: 10.1007/s12192-010-0190-1. Cell Stress Chaperones. 2010. PMID: 20393890 Free PMC article.
-
Scarless integumentary wound healing in the mammalian fetus: molecular basis and therapeutic implications.Birth Defects Res C Embryo Today. 2012 Sep;96(3):223-36. doi: 10.1002/bdrc.21015. Birth Defects Res C Embryo Today. 2012. PMID: 23109318 Review.
-
[The modern approach to wound treatment].Med Pregl. 2000 Jul-Aug;53(7-8):363-8. Med Pregl. 2000. PMID: 11214479 Review. Croatian.
Cited by
-
Hyaluronic acid-curcumin conjugate suppresses the fibrotic functions of myofibroblasts from contractive joint by the PTGER2 demethylation.Regen Biomater. 2019 Oct;6(5):269-277. doi: 10.1093/rb/rbz016. Epub 2019 Apr 22. Regen Biomater. 2019. PMID: 31616564 Free PMC article.
-
Cellular and Molecular Characteristics of Scarless versus Fibrotic Wound Healing.Dermatol Res Pract. 2010;2010:790234. doi: 10.1155/2010/790234. Epub 2010 Dec 27. Dermatol Res Pract. 2010. PMID: 21253544 Free PMC article.
-
Overexpression of chaperonin containing T-complex polypeptide subunit zeta 2 (CCT6b) suppresses the functions of active fibroblasts in a rat model of joint contracture.J Orthop Surg Res. 2019 May 9;14(1):125. doi: 10.1186/s13018-019-1161-6. J Orthop Surg Res. 2019. PMID: 31072365 Free PMC article.
-
Identification and functional analysis of healing regulators in Drosophila.PLoS Genet. 2015 Feb 3;11(2):e1004965. doi: 10.1371/journal.pgen.1004965. eCollection 2015. PLoS Genet. 2015. PMID: 25647511 Free PMC article.
-
Chaperonin containing T-complex polypeptide (CCT) subunit expression in oral mucosal wounds and fibroblasts.Cell Stress Chaperones. 2011 Nov;16(6):675-80. doi: 10.1007/s12192-011-0274-6. Epub 2011 Jun 28. Cell Stress Chaperones. 2011. PMID: 21710295 Free PMC article.
References
-
- Clark RAF. Wound Repair: overview and general considerations. In: Clark RAF, editor. The molecular and cellular biology of wound repair. 2nd ed. New York: Plenum Press; 1996. pp. 3–50.
-
- Hantash BM, Zhao L, Knowles JA, Lorenz PH. Adult and fetal wound healing. Front Biosci. 2008;13:51–61. - PubMed
-
- Colwell AS, Longaker MT, Lorenz PH. Mammalian fetal organ regeneration. Adv Biochem Eng Biotechnol. 2005;93:83–100. - PubMed
-
- Wilgus TA. Regenerative healing in fetal skin: A review of the literature. Ostomy Wound Manage. 2007;53:16–31. - PubMed
-
- Gabbiani G. The myofibroblast in wound healing and fibrocontractive diseases. J Pathol. 2003;200:500–503. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
