

Functional and genetic studies of the substrate specificity of coronavirus infectious bronchitis virus 3C-like proteinase
- PMID: 20444893
- PMCID: PMC2898227
- DOI: 10.1128/JVI.02490-09
Functional and genetic studies of the substrate specificity of coronavirus infectious bronchitis virus 3C-like proteinase
Abstract
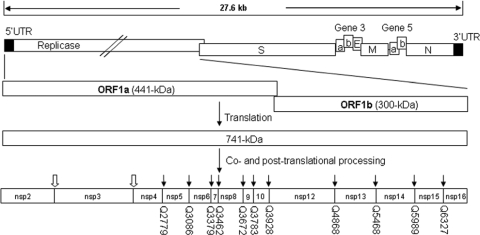
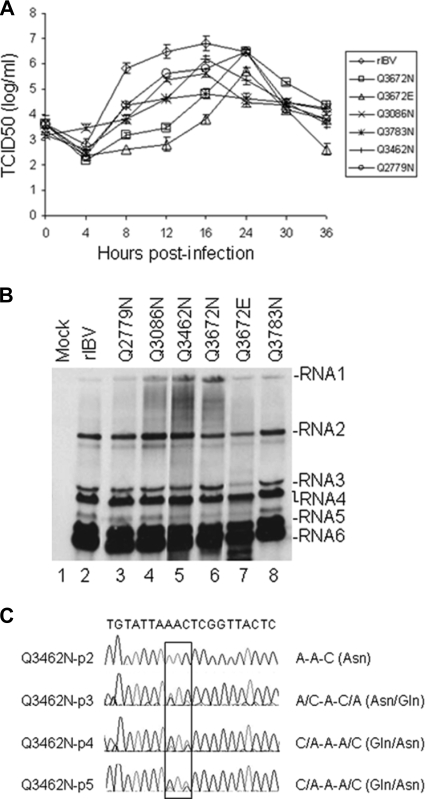
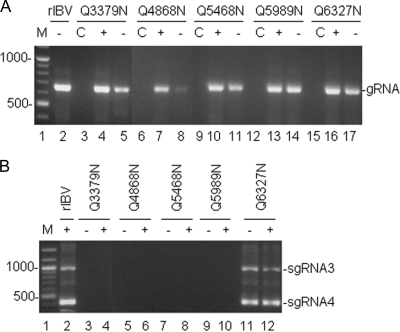
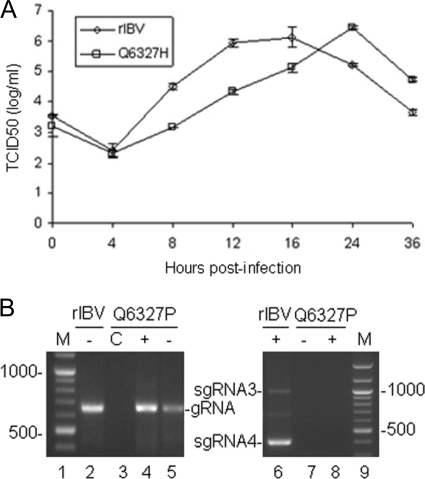
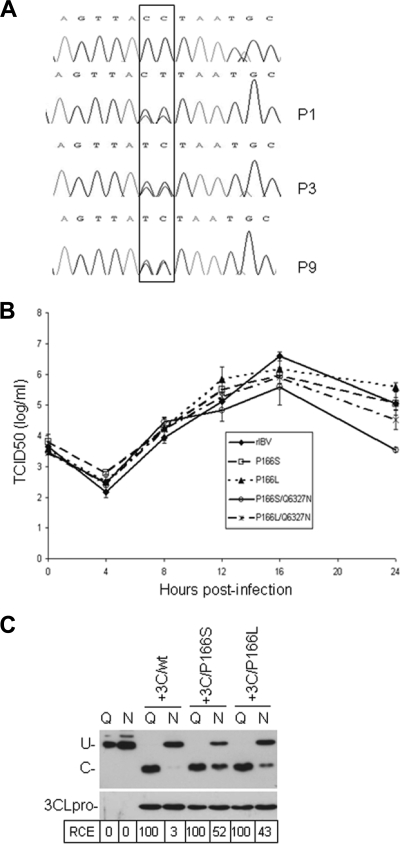
Coronavirus (CoV) 3C-like proteinase (3CLpro), located in nonstructural protein 5 (nsp5), processes the replicase polyproteins 1a and 1ab (pp1a and pp1ab) at 11 specific sites to produce 12 mature nonstructural proteins (nsp5 to nsp16). Structural and biochemical studies suggest that a conserved Gln residue at the P1 position is absolutely required for efficient cleavage. Here, we investigate the effects of amino acid substitution at the P1 position of 3CLpro cleavage sites of infectious bronchitis virus (IBV) on the cleavage efficiency and viral replication by in vitro cleavage assays and reverse genetic approaches. Our results demonstrated that a P1-Asn substitution at the nsp4-5/Q2779, nsp5-6/Q3086, nsp7-8/Q3462, nsp8-9/Q3672, and nsp9-10/Q3783 sites, a P1-Glu substitution at the nsp8-9/Q3672 site, and a P1-His substitution at the nsp15-16/Q6327 site were tolerated and allowed recovery of infectious mutant viruses, albeit with variable degrees of growth defects. In contrast, a P1-Asn substitution at the nsp6-7/Q3379, nsp12-13/Q4868, nsp13-14/Q5468, and nsp14-15/Q5989 sites, as well as a P1-Pro substitution at the nsp15-16/Q6327 site, abolished 3CLpro-mediated cleavage at the corresponding position and blocked the recovery of infectious viruses. Analysis of the effects of these lethal mutations on RNA synthesis suggested that processing intermediates, such as the nsp6-7, nsp12-13, nsp13-14, nsp14-15, and nsp15-16 precursors, may function in negative-stranded genomic RNA replication, whereas mature proteins may be required for subgenomic RNA (sgRNA) transcription. More interestingly, a mutant 3CLpro with either a P166S or P166L mutation was selected when an IBV infectious cDNA clone carrying the Q6327N mutation at the nsp15-16 site was introduced into cells. Either of the two mutations was proved to enhance significantly the 3CLpro-mediated cleavage efficiency at the nsp15-16 site with a P1-Asn substitution and compensate for the detrimental effects on recovery of infectious virus.
Figures

Similar articles
-
Proteolytic processing of polyproteins 1a and 1ab between non-structural proteins 10 and 11/12 of Coronavirus infectious bronchitis virus is dispensable for viral replication in cultured cells.Virology. 2008 Sep 30;379(2):175-80. doi: 10.1016/j.virol.2008.06.038. Epub 2008 Aug 3. Virology. 2008. PMID: 18678384 Free PMC article.
-
Further characterization of the coronavirus infectious bronchitis virus 3C-like proteinase and determination of a new cleavage site.Virology. 2000 Jun 20;272(1):27-39. doi: 10.1006/viro.2000.0330. Virology. 2000. PMID: 10873746 Free PMC article.
-
Further requirements for cleavage by the murine coronavirus 3C-like proteinase: identification of a cleavage site within ORF1b.Virology. 1999 Oct 25;263(2):471-84. doi: 10.1006/viro.1999.9954. Virology. 1999. PMID: 10544119 Free PMC article.
-
Potential 3-chymotrypsin-like cysteine protease cleavage sites in the coronavirus polyproteins pp1a and pp1ab and their possible relevance to COVID-19 vaccine and drug development.FASEB J. 2021 May;35(5):e21573. doi: 10.1096/fj.202100280RR. FASEB J. 2021. PMID: 33913206 Free PMC article. Review.
-
Current Knowledge on Infectious Bronchitis Virus Non-structural Proteins: The Bearer for Achieving Immune Evasion Function.Front Vet Sci. 2022 Apr 8;9:820625. doi: 10.3389/fvets.2022.820625. eCollection 2022. Front Vet Sci. 2022. PMID: 35464391 Free PMC article. Review.
Cited by
-
A Brief Review: The Z-curve Theory and its Application in Genome Analysis.Curr Genomics. 2014 Apr;15(2):78-94. doi: 10.2174/1389202915999140328162433. Curr Genomics. 2014. PMID: 24822026 Free PMC article.
-
The main protease (Mpro) from SARS-CoV-2 triggers plasma clotting in vitro by activating coagulation factors VII and FXII.Commun Biol. 2025 Aug 1;8(1):1145. doi: 10.1038/s42003-025-08570-2. Commun Biol. 2025. PMID: 40750828 Free PMC article.
-
Temperature-sensitive mutants and revertants in the coronavirus nonstructural protein 5 protease (3CLpro) define residues involved in long-distance communication and regulation of protease activity.J Virol. 2012 May;86(9):4801-10. doi: 10.1128/JVI.06754-11. Epub 2012 Feb 15. J Virol. 2012. PMID: 22345451 Free PMC article.
-
Characterization of Bafinivirus main protease autoprocessing activities.J Virol. 2011 Feb;85(3):1348-59. doi: 10.1128/JVI.01716-10. Epub 2010 Nov 10. J Virol. 2011. PMID: 21068254 Free PMC article.
-
Host-specific sensing of coronaviruses and picornaviruses by the CARD8 inflammasome.PLoS Biol. 2023 Jun 8;21(6):e3002144. doi: 10.1371/journal.pbio.3002144. eCollection 2023 Jun. PLoS Biol. 2023. PMID: 37289745 Free PMC article.
References
-
- Almazán, F., M. L. Dediego, C. Galan, D. Escors, E. Alvarez, J. Ortego, I. Sola, S. Zuniga, S. Alonso, J. L. Moreno, A. Nogales, C. Capiscol, and L. Enjuanes. 2006. Construction of a severe acute respiratory syndrome coronavirus infectious cDNA clone and a replicon to study coronavirus RNA synthesis. J. Virol. 80:10900-10906. - PMC - PubMed
-
- Anand, K., J. Ziebuhr, P. Wadhwani, J. R. Mesters, and R. Hilgenfeld. 2003. Coronavirus main proteinase (3CLpro) structure: basis for design of anti-SARS drugs. Science 300:1763-1767. - PubMed
-
- Barrette-Ng, I. H., K. K. Ng, B. L. Mark, D. Van Aken, M. M. Cherney, C. Garen, Y. Kolodenko, A. E. Gorbalenya, E. J. Snijder, and M. N. James. 2002. Structure of arterivirus nsp4. The smallest chymotrypsin-like proteinase with an alpha/beta C-terminal extension and alternate conformations of the oxyanion hole. J. Biol. Chem. 277:39960-39966. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
