

Recovery of heat shock-triggered released apoplastic Ca2+ accompanied by pectin methylesterase activity is required for thermotolerance in soybean seedlings
- PMID: 20444907
- PMCID: PMC2882276
- DOI: 10.1093/jxb/erq121
Recovery of heat shock-triggered released apoplastic Ca2+ accompanied by pectin methylesterase activity is required for thermotolerance in soybean seedlings
Abstract
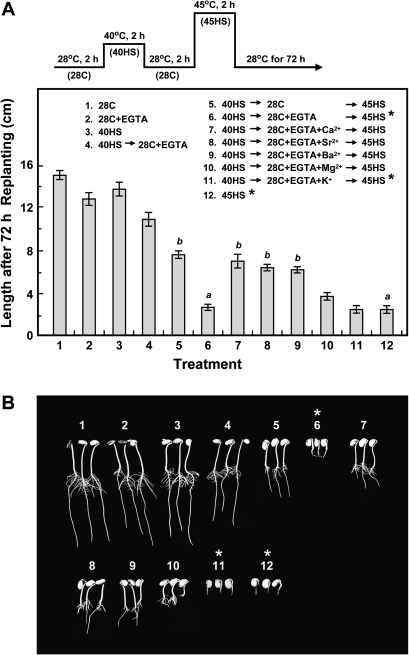
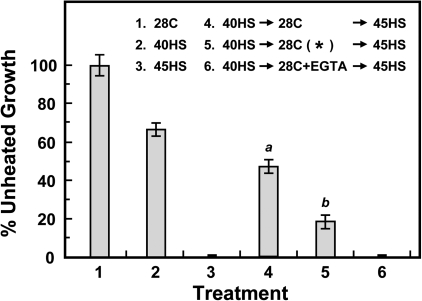
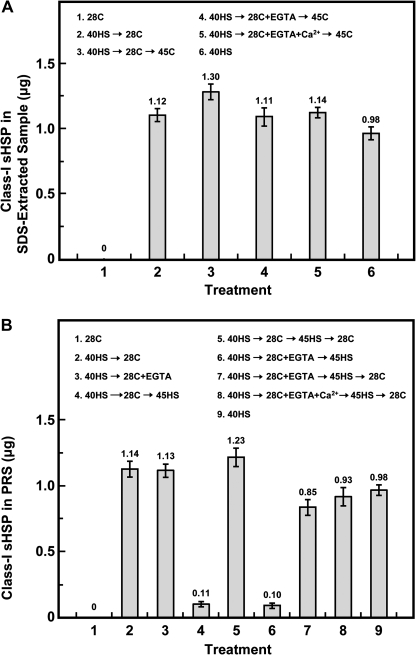
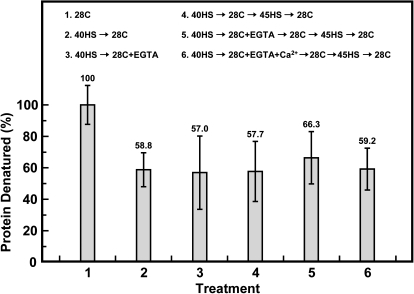
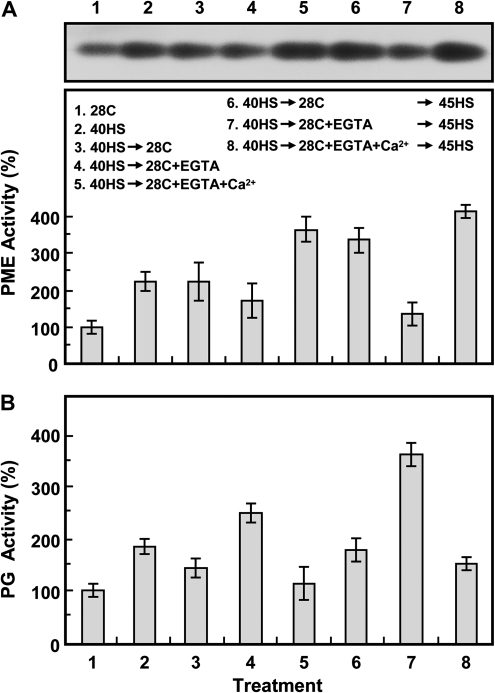
Synthesis of heat shock proteins (HSPs) in response to heat shock (HS) is essential for thermotolerance. The effect of a Ca(2+) chelator, EGTA, was investigated before a lethal HS treatment in soybean (Glycine max) seedlings with acquired thermotolerance induced by preheating. Such seedlings became non-thermotolerant with EGTA treatment. The addition of Ca(2+), Sr(2+) or Ba(2+) to the EGTA-treated samples rescued the seedlings from death by preventing the increased cellular leakage of electrolytes, amino acids, and sugars caused by EGTA. It was confirmed that EGTA did not affect HSP accumulation and physiological functions but interfered with the recovery of HS-released Ca(2+) concentration which was required for thermotolerance. Pectin methylesterase (PME, EC 3.1.1.11), a cell wall remodelling enzyme, was activated in response to HS, and its elevated activity caused an increased level of demethylesterified pectin which was related to the recovery of the HS-released Ca(2+) concentration. Thus, the recovery of HS-released Ca(2+) in Ca(2+)-pectate reconstitution through PME activity is required for cell wall remodelling during HS in soybean which, in turn, retains plasma membrane integrity and co-ordinates with HSPs to confer thermotolerance.
Figures




Similar articles
-
Heat shock-triggered Ca2+ mobilization accompanied by pectin methylesterase activity and cytosolic Ca2+ oscillation are crucial for plant thermotolerance.Plant Signal Behav. 2010 Oct;5(10):1252-6. doi: 10.4161/psb.5.10.12607. Epub 2010 Oct 1. Plant Signal Behav. 2010. PMID: 20948293 Free PMC article.
-
Crosstalk of methylglyoxal and calcium signaling in maize (Zea mays L.) thermotolerance through methylglyoxal-scavenging system.J Plant Physiol. 2024 Dec;303:154362. doi: 10.1016/j.jplph.2024.154362. Epub 2024 Oct 4. J Plant Physiol. 2024. PMID: 39395220
-
Cyclic nucleotide-gated ion channel 6 mediates thermotolerance in Arabidopsis seedlings by regulating nitric oxide production via cytosolic calcium ions.BMC Plant Biol. 2019 Aug 20;19(1):368. doi: 10.1186/s12870-019-1974-9. BMC Plant Biol. 2019. PMID: 31429706 Free PMC article.
-
Pectin methylesterase is required for guard cell function in response to heat.Plant Signal Behav. 2017 Jun 3;12(6):e1338227. doi: 10.1080/15592324.2017.1338227. Epub 2017 Jun 15. Plant Signal Behav. 2017. PMID: 28617153 Free PMC article. Review.
-
Thermotolerance and the heat shock proteins.Symp Soc Exp Biol. 1987;41:269-83. Symp Soc Exp Biol. 1987. PMID: 3332487 Review.
Cited by
-
Pectin Methylesterases: Cell Wall Remodeling Proteins Are Required for Plant Response to Heat Stress.Front Plant Sci. 2018 Nov 6;9:1612. doi: 10.3389/fpls.2018.01612. eCollection 2018. Front Plant Sci. 2018. PMID: 30459794 Free PMC article. Review.
-
Guard Cell-Specific Pectin METHYLESTERASE53 Is Required for Abscisic Acid-Mediated Stomatal Function and Heat Response in Arabidopsis.Front Plant Sci. 2022 Feb 21;13:836151. doi: 10.3389/fpls.2022.836151. eCollection 2022. Front Plant Sci. 2022. PMID: 35265095 Free PMC article.
-
Cell wall, lignin and fatty acid-related transcriptome in soybean: Achieving gene expression patterns for bioenergy legume.Genet Mol Biol. 2012 Jun;35(1 (suppl)):322-30. doi: 10.1590/S1415-47572012000200013. Genet Mol Biol. 2012. PMID: 22802717 Free PMC article.
-
Heat shock-triggered Ca2+ mobilization accompanied by pectin methylesterase activity and cytosolic Ca2+ oscillation are crucial for plant thermotolerance.Plant Signal Behav. 2010 Oct;5(10):1252-6. doi: 10.4161/psb.5.10.12607. Epub 2010 Oct 1. Plant Signal Behav. 2010. PMID: 20948293 Free PMC article.
-
BIIDXI, a DUF642 Cell Wall Protein That Regulates Pectin Methyl Esterase Activity, Is Involved in Thermotolerance Processes in Arabidopsis thaliana.Plants (Basel). 2022 Nov 11;11(22):3049. doi: 10.3390/plants11223049. Plants (Basel). 2022. PMID: 36432778 Free PMC article.
References
-
- Alexandrov V. Functional aspects of cell response to heat shock. International Review of Cytology. 1994;148:171–227. - PubMed
-
- Crelier S, Robert M-C, Claude J, Juillerat M-A. Tomato (Lycopersicon esculentum) pectin methylesterase and polygalacturonase behaviors regarding heat- and pressure-induced inactivation. Journal of Agricultural and Food Chemistry. 2001;49:5566–5575. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
