

Memory impairment in transgenic Alzheimer mice requires cellular prion protein
- PMID: 20445063
- PMCID: PMC3323924
- DOI: 10.1523/JNEUROSCI.0395-10.2010
Memory impairment in transgenic Alzheimer mice requires cellular prion protein
Abstract
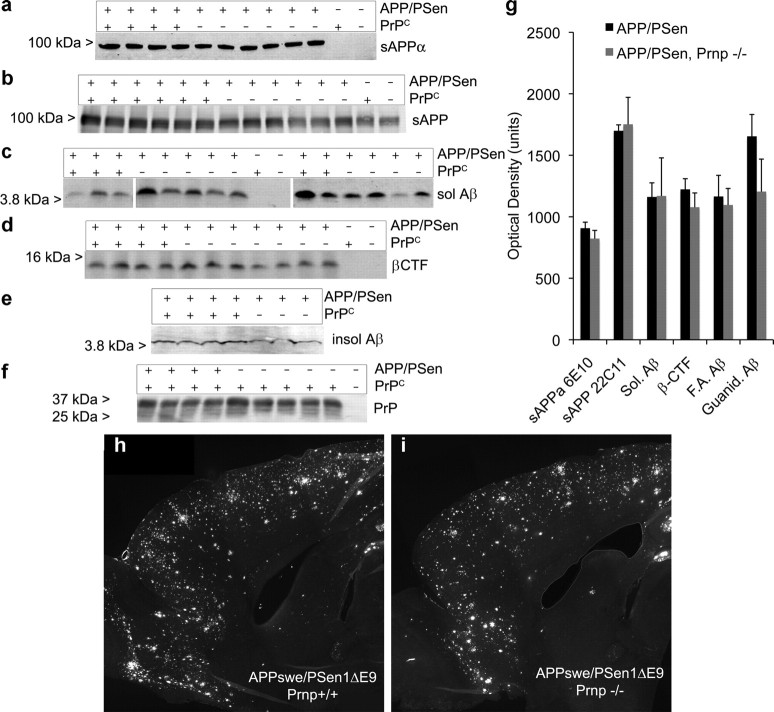
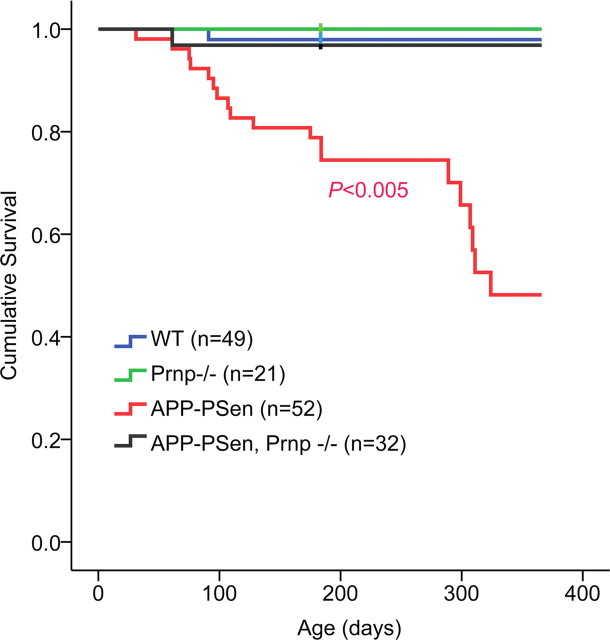
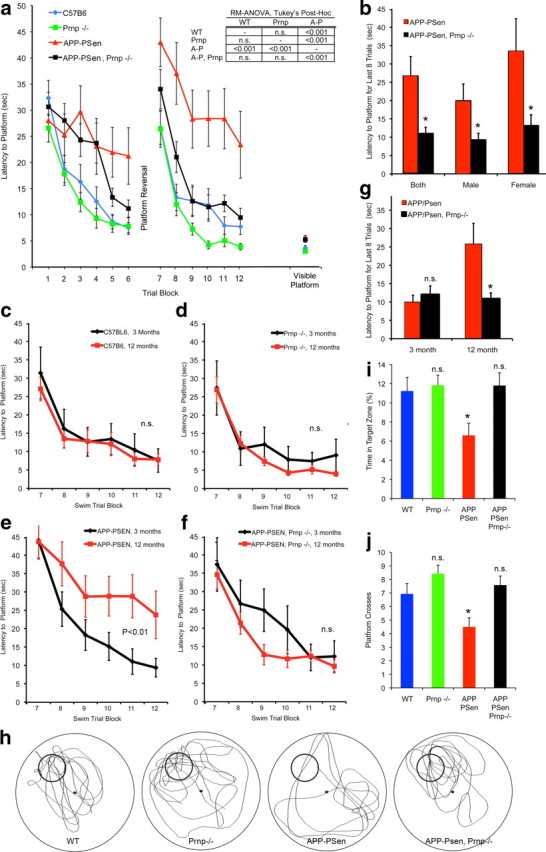
Soluble oligomers of the amyloid-beta (Abeta) peptide are thought to play a key role in the pathophysiology of Alzheimer's disease (AD). Recently, we reported that synthetic Abeta oligomers bind to cellular prion protein (PrP(C)) and that this interaction is required for suppression of synaptic plasticity in hippocampal slices by oligomeric Abeta peptide. We hypothesized that PrP(C) is essential for the ability of brain-derived Abeta to suppress cognitive function. Here, we crossed familial AD transgenes encoding APPswe and PSen1DeltaE9 into Prnp-/- mice to examine the necessity of PrP(C) for AD-related phenotypes. Neither APP expression nor Abeta level is altered by PrP(C) absence in this transgenic AD model, and astrogliosis is unchanged. However, deletion of PrP(C) expression rescues 5-HT axonal degeneration, loss of synaptic markers, and early death in APPswe/PSen1DeltaE9 transgenic mice. The AD transgenic mice with intact PrP(C) expression exhibit deficits in spatial learning and memory. Mice lacking PrP(C), but containing Abeta plaque derived from APPswe/PSen1DeltaE9 transgenes, show no detectable impairment of spatial learning and memory. Thus, deletion of PrP(C) expression dissociates Abeta accumulation from behavioral impairment in these AD mice, with the cognitive deficits selectively requiring PrP(C).
Figures

Comment in
-
Is PrPC a mediator of Abeta toxicity in Alzheimer's disease?J Neurosci. 2010 Sep 8;30(36):11883-4. doi: 10.1523/JNEUROSCI.3235-10.2010. J Neurosci. 2010. PMID: 20826652 Free PMC article. No abstract available.
References
-
- Austin MC, Whitehead RE, Edgar CL, Janosky JE, Lewis DA. Localized decrease in serotonin transporter-immunoreactive axons in the prefrontal cortex of depressed subjects committing suicide. Neuroscience. 2002;114:807–815. - PubMed
-
- Balducci C, Beeg M, Stravalaci M, Bastone A, Sclip A, Biasini E, Tapella L, Colombo L, Manzoni C, Borsello T, Chiesa R, Gobbi M, Salmona M, Forloni G. Synthetic amyloid-beta oligomers impair long-term memory independently of cellular prion protein. Proc Natl Acad Sci U S A. 2010;107:2295–2300. - PMC - PubMed
-
- Berlyne DE. Curiosity and exploration. Science. 1966;153:25–33. - PubMed
-
- Berlyne DE, Koenig ID, Hirota T. Novelty, arousal, and the reinforcement of diversive exploration in the rat. J Comp Physiol Psychol. 1966;62:222–226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials