

Formation of oral and pharyngeal dentition in teleosts depends on differential recruitment of retinoic acid signaling
- PMID: 20445074
- PMCID: PMC2923347
- DOI: 10.1096/fj.09-147488
Formation of oral and pharyngeal dentition in teleosts depends on differential recruitment of retinoic acid signaling
Abstract
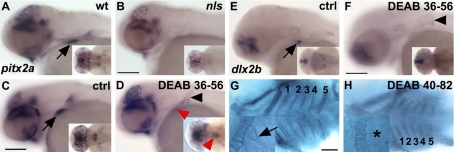
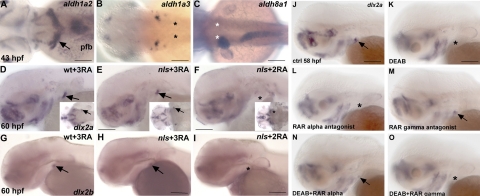
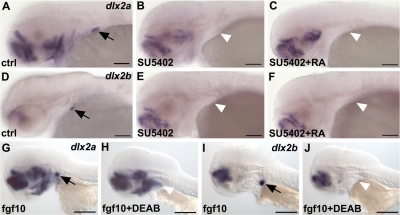
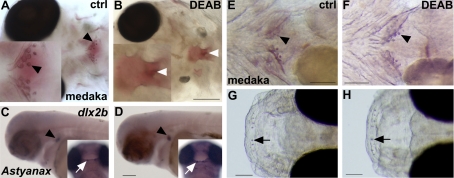
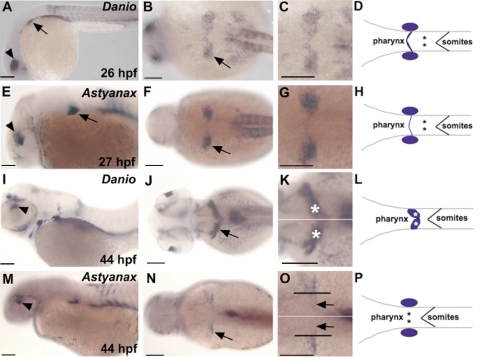
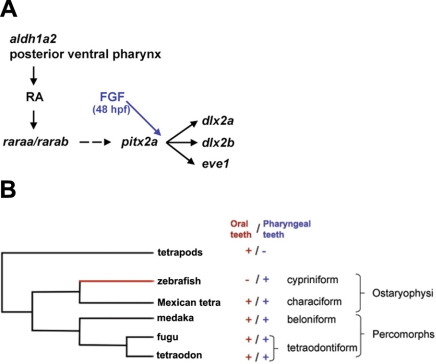
One of the goals of evolutionary developmental biology is to link specific adaptations to changes in developmental pathways. The dentition of cypriniform fishes, which in contrast to many other teleost fish species possess pharyngeal teeth but lack oral teeth, provides a suitable model to study the development of feeding adaptations. Here, we have examined the involvement of retinoic acid (RA) in tooth development and show that RA is specifically required to induce the pharyngeal tooth developmental program in zebrafish. Perturbation of RA signaling at this stage abolished tooth induction without affecting the development of tooth-associated ceratobranchial bones. We show that this inductive event is dependent on RA synthesis from aldh1a2 in the ventral posterior pharynx. Fibroblast growth factor (FGF) signaling has been shown to be critical for tooth induction in zebrafish, and its loss has been associated with oral tooth loss in cypriniform fishes. Pharmacological treatments targeting the RA and FGF pathways revealed that both pathways act independently during tooth induction. In contrast, we find that in Mexican tetra and medaka, species that also possess oral teeth, both oral and pharyngeal teeth are induced independently of RA. Our analyses suggest an evolutionary scenario in which the gene network controlling tooth development obtained RA dependency in the lineage leading to the cypriniforms. The loss of pharyngeal teeth in this group was cancelled out through a shift in aldh1a2 expression, while oral teeth might have been lost ultimately due to deficient RA signaling in the oral cavity.
Figures
References
-
- Smith M M, Fraser G J, Mitsiadis T A. Dental lamina as source of odontogenic stem cells: evolutionary origins and developmental control of tooth generation in gnathostomes. J Exp Zoolog B Mol Dev Evol. 2009;312B:260–280. - PubMed
-
- Miletich I, Sharpe P T. Neural crest contribution to mammalian tooth formation. Birth Defects Res C Embryo Today. 2004;72:200–212. - PubMed
-
- Mitsiadis T A, Smith M M. How do genes make teeth to order through development? J Exp Zoolog B Mol Dev Evol. 2006;306:177–182. - PubMed
-
- Tompkins K. Molecular mechanisms of cytodifferentiation in mammalian tooth development. Connect Tissue Res. 2006;47:111–118. - PubMed
-
- Stock D W. Zebrafish dentition in comparative context. J Exp Zoolog B Mol Dev Evol. 2007;308:523–549. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
