

Biophysical properties of mitochondrial fusion events in pancreatic beta-cells and cardiac cells unravel potential control mechanisms of its selectivity
- PMID: 20445168
- PMCID: PMC2928626
- DOI: 10.1152/ajpcell.00427.2009
Biophysical properties of mitochondrial fusion events in pancreatic beta-cells and cardiac cells unravel potential control mechanisms of its selectivity
Abstract
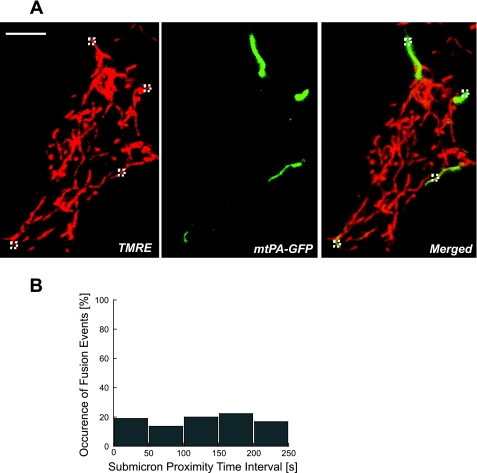
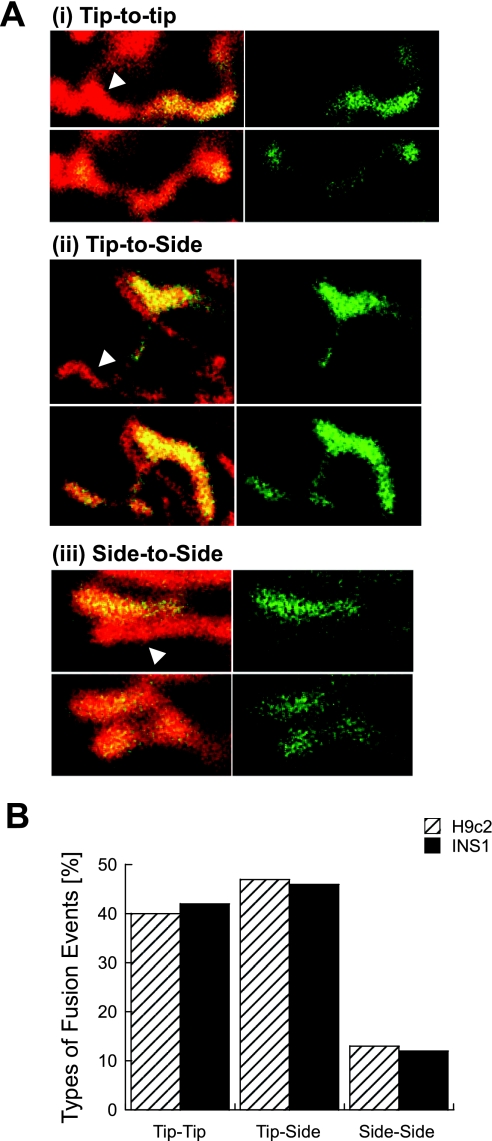
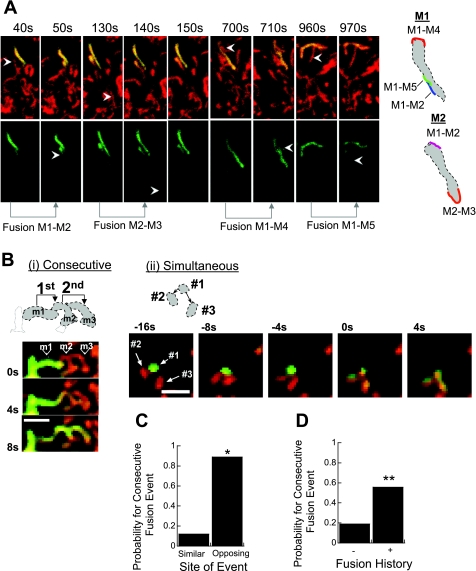
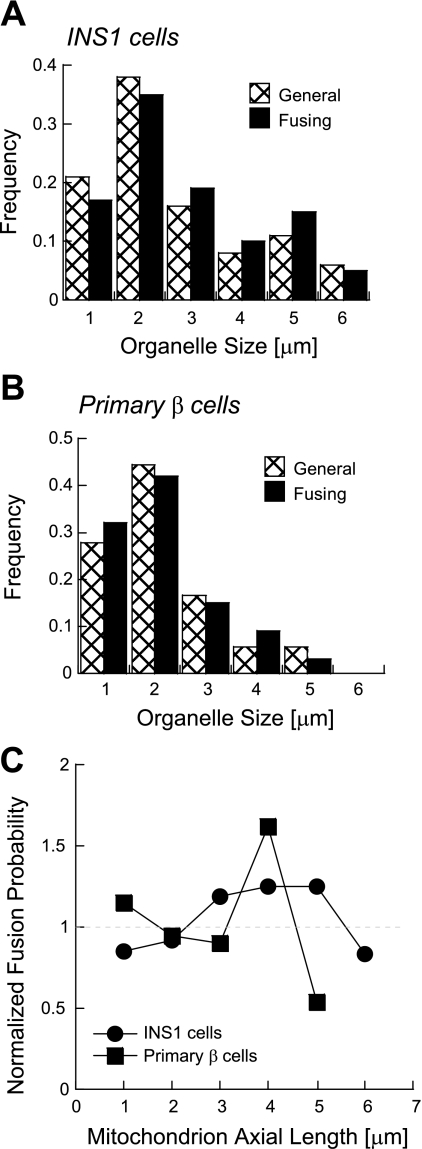
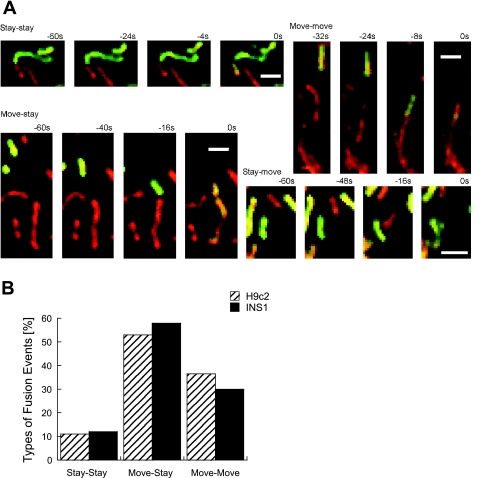
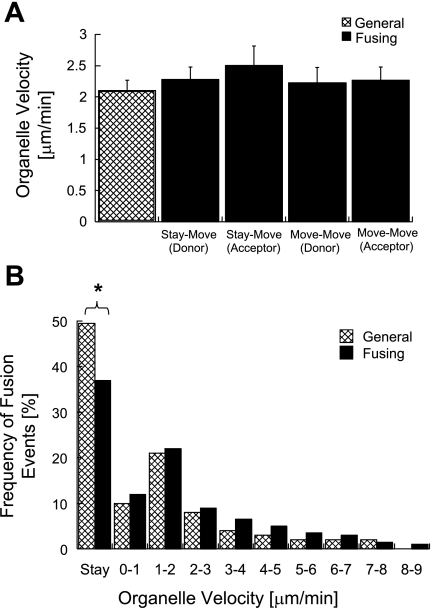
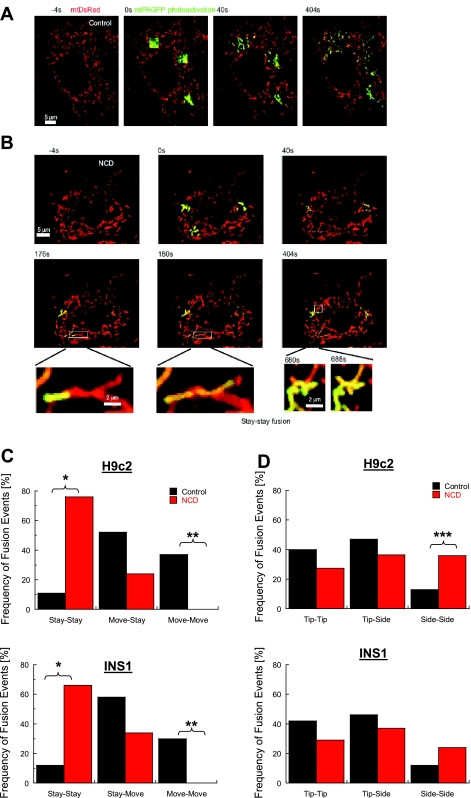
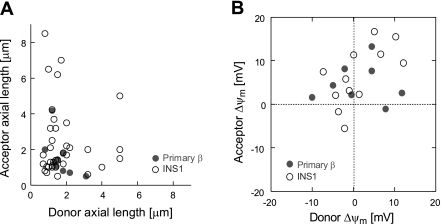
Studies in various types of cells find that, on average, each mitochondrion becomes involved in a fusion event every 15 min, depending on the cell type. As most contact events do not result in mitochondrial fusion, it is expected that properties of the individual mitochondrion determine the likelihood of a fusion event. However, apart from membrane potential, the properties that influence the likelihood of entering a fusion event are not known. Here, we tag and track individual mitochondria in H9c2, INS1, and primary beta-cells and determine the biophysical properties that increase the likelihood of a fusion event. We found that the probability for fusion is independent of contact duration and organelle dimensions, but it is influenced by organelle motility. Furthermore, the history of a previous fusion event of the individual mitochondrion influenced both the likelihood for a subsequent fusion event, as well as the site on the mitochondrion at which the fusion occurred. These observations unravel the specific properties that distinguish mitochondria that will enter fusion events from the ones that will not. Altogether, these properties may help to elucidate the molecular mechanisms that regulate fusion at the level of the single mitochondrion.
Figures
References
-
- Bindokas VP, Kuznetsov A, Sreenan S, Polonsky KS, Roe MW, Philipson LH. Visualizing superoxide production in normal and diabetic rat islets of Langerhans. J Biol Chem 278: 9796–9801, 2003 - PubMed
-
- Busch KB, Bereiter-Hahn J, Wittig I, Schagger H, Jendrach M. Mitochondrial dynamics generate equal distribution but patchwork localization of respiratory. Complex I Mol Membr Biol 23: 509–520, 2006 - PubMed
-
- Chada SR, Hollenbeck PJ. Mitochondrial movement and positioning in axons: the role of growth factor signaling. J Exp Biol 206: 1985–1992, 2003 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials