

On the induction of postsynaptic granule cell-Purkinje neuron LTP and LTD
- PMID: 20446074
- PMCID: PMC3103506
- DOI: 10.1007/s12311-010-0174-9
On the induction of postsynaptic granule cell-Purkinje neuron LTP and LTD
Abstract
In the last decade, several experimental studies have demonstrated that particular patterns of synaptic activity can induce postsynaptic parallel fiber (PF) long-term potentiation (LTP). This form of plasticity can reverse postsynaptic PF long-term depression (LTD), which has been traditionally considered as the principal form of plasticity underlying cerebellar learning. Postsynaptic PF-LTP requires a transient increase in intracellular Ca(2+) concentration and, in contrast to PF-LTD, is induced without concomitant climbing fiber (CF) activation. Thus, it has been postulated that the polarity of long-term synaptic plasticity is determined by the amplitude of the Ca(2+) transient during the induction protocol, with PF-LTP induced by smaller Ca(2+) signals without concomitant CF activation. However, this hypothesis is contradicted by recent studies. A quantitative analysis of Ca(2+) signals associated with induction of PF-LTP indicates that the bidirectional induction of long-term plasticity is regulated by more complex mechanisms. Here we review the state-of-the-art of research on postsynaptic PF-LTP and PF-LTD and discuss the principal open questions on this topic.
Figures
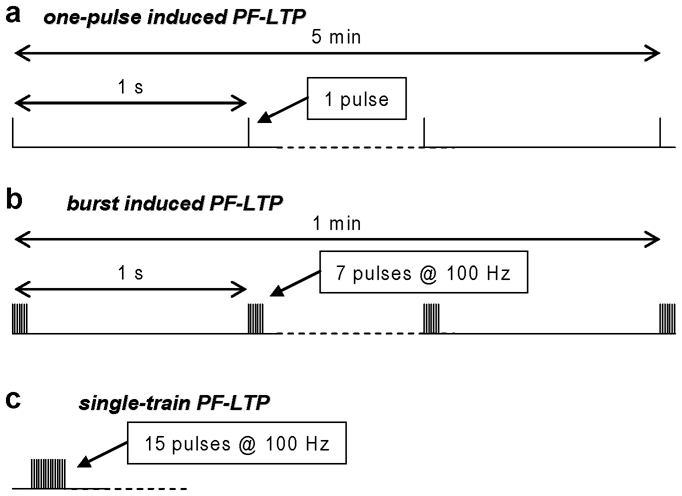
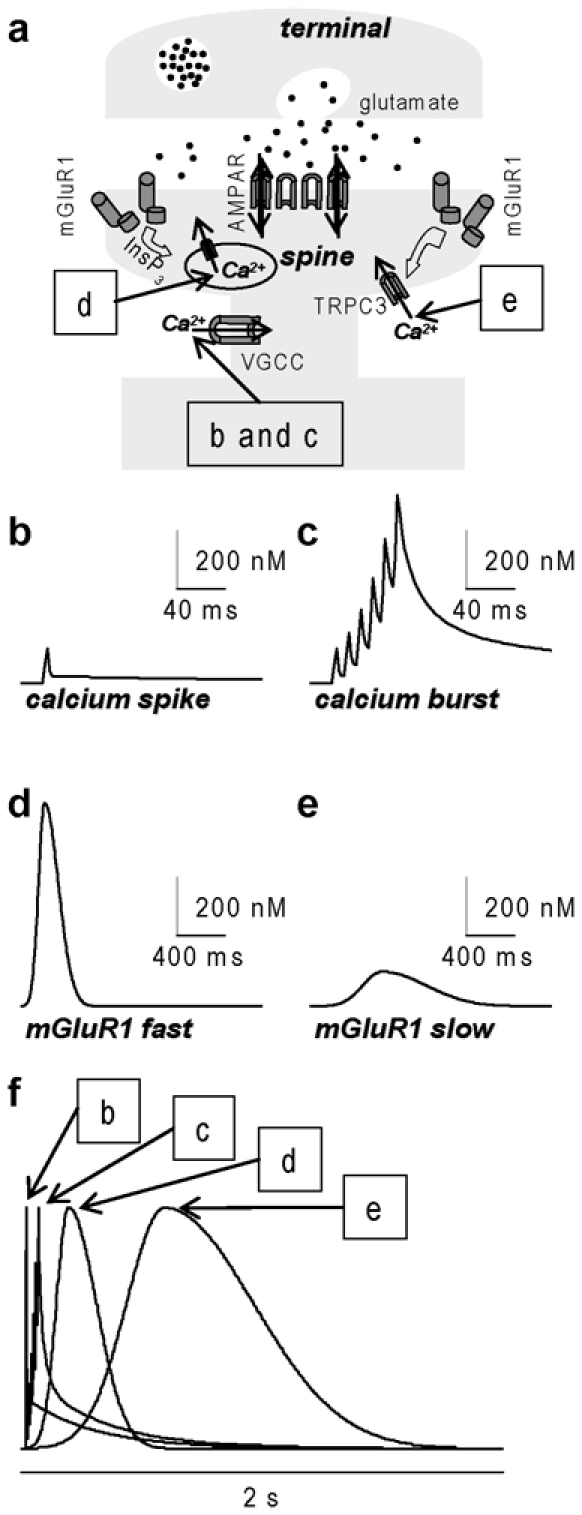
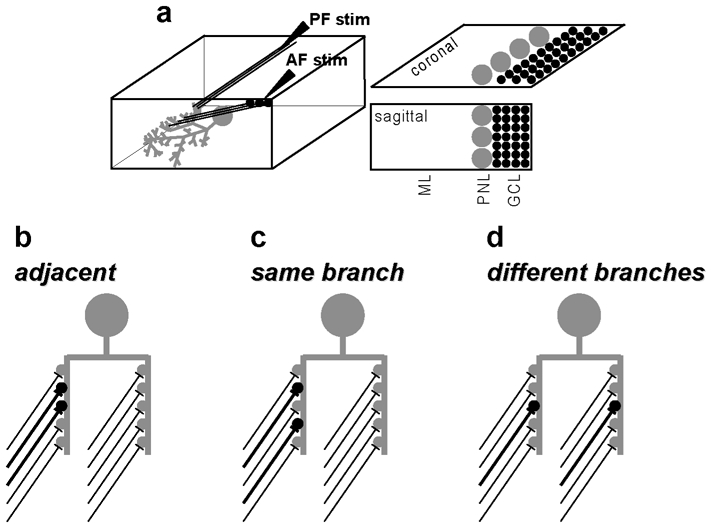
Similar articles
-
Long-Term Depression of Intrinsic Excitability Accompanied by Synaptic Depression in Cerebellar Purkinje Cells.J Neurosci. 2017 Jun 7;37(23):5659-5669. doi: 10.1523/JNEUROSCI.3464-16.2017. Epub 2017 May 11. J Neurosci. 2017. PMID: 28495974 Free PMC article.
-
Different Numbers of Conjunctive Stimuli Induce LTP or LTD in Mouse Cerebellar Purkinje Cell.Cerebellum. 2024 Dec;23(6):2297-2307. doi: 10.1007/s12311-024-01726-6. Epub 2024 Aug 3. Cerebellum. 2024. PMID: 39096432 Free PMC article.
-
Cell type-specific plasticity at parallel fiber synapses onto Purkinje cells in the posterior caudal lobe of the mormyrid fish cerebellum.J Neurophysiol. 2018 Aug 1;120(2):644-661. doi: 10.1152/jn.00175.2018. Epub 2018 Apr 18. J Neurophysiol. 2018. PMID: 29668384
-
Synaptic memories upside down: bidirectional plasticity at cerebellar parallel fiber-Purkinje cell synapses.Neuron. 2006 Oct 19;52(2):227-38. doi: 10.1016/j.neuron.2006.09.032. Neuron. 2006. PMID: 17046686 Review.
-
Cerebellar long-term potentiation: cellular mechanisms and role in learning.Int Rev Neurobiol. 2014;117:39-51. doi: 10.1016/B978-0-12-420247-4.00003-8. Int Rev Neurobiol. 2014. PMID: 25172628 Review.
Cited by
-
Reactive Oxygen Species: Physiological and Physiopathological Effects on Synaptic Plasticity.J Exp Neurosci. 2016 Sep 4;10(Suppl 1):23-48. doi: 10.4137/JEN.S39887. eCollection 2016. J Exp Neurosci. 2016. PMID: 27625575 Free PMC article. Review.
-
Number of spikes in climbing fibers determines the direction of cerebellar learning.J Neurosci. 2013 Aug 14;33(33):13436-40. doi: 10.1523/JNEUROSCI.1527-13.2013. J Neurosci. 2013. PMID: 23946401 Free PMC article.
-
A stochastic signaling network mediates the probabilistic induction of cerebellar long-term depression.J Neurosci. 2012 Jul 4;32(27):9288-300. doi: 10.1523/JNEUROSCI.5976-11.2012. J Neurosci. 2012. PMID: 22764236 Free PMC article.
-
De Novo Synthesized Estradiol: A Role in Modulating the Cerebellar Function.Int J Mol Sci. 2020 May 7;21(9):3316. doi: 10.3390/ijms21093316. Int J Mol Sci. 2020. PMID: 32392845 Free PMC article. Review.
-
Two Distinct Sets of Ca2+ and K+ Channels Are Activated at Different Membrane Potentials by the Climbing Fiber Synaptic Potential in Purkinje Neuron Dendrites.J Neurosci. 2019 Mar 13;39(11):1969-1981. doi: 10.1523/JNEUROSCI.2155-18.2018. Epub 2019 Jan 10. J Neurosci. 2019. PMID: 30630881 Free PMC article.
References
-
- Albus JS. A theory of cerebellar function. Math Biosci. 1971;28:167–171.
-
- Karachot L, Kado RT, Ito M. Stimulus parameters for induction of long-term depression in in vitro rat Purkinje cells. Neurosci Res. 1995;21:161–168. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
