

The tertiary structure of group II introns: implications for biological function and evolution
- PMID: 20446804
- PMCID: PMC4408542
- DOI: 10.3109/10409231003796523
The tertiary structure of group II introns: implications for biological function and evolution
Abstract
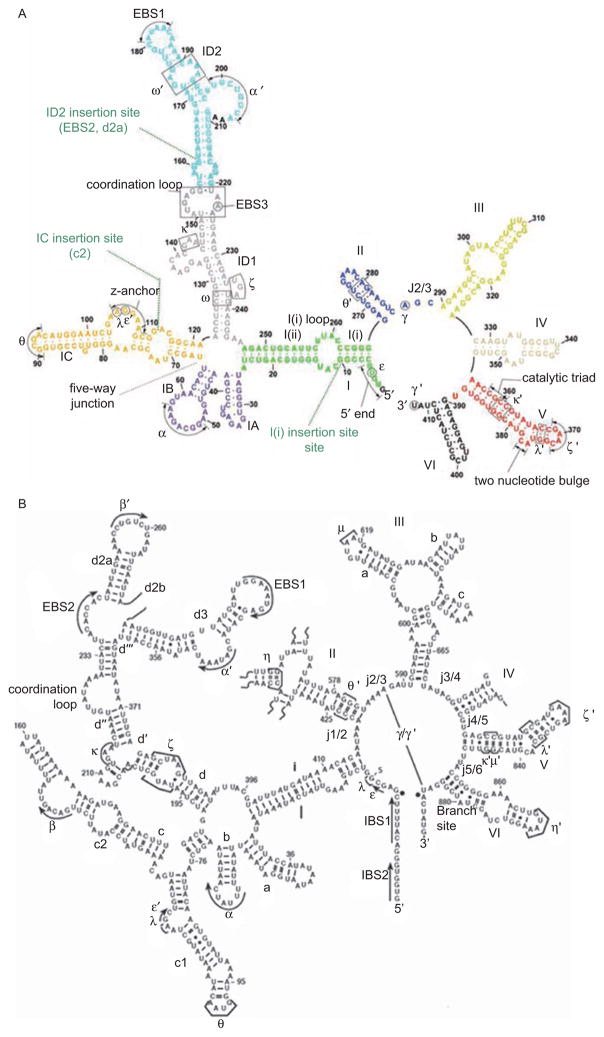
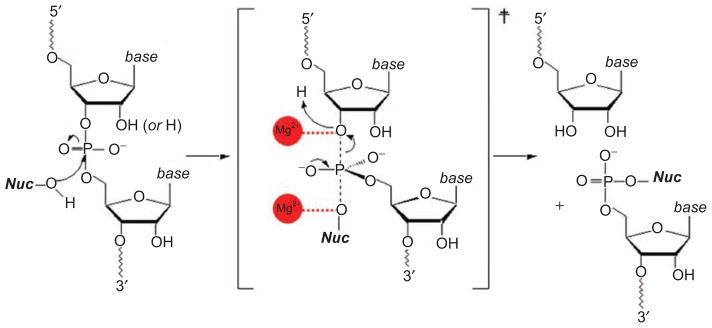
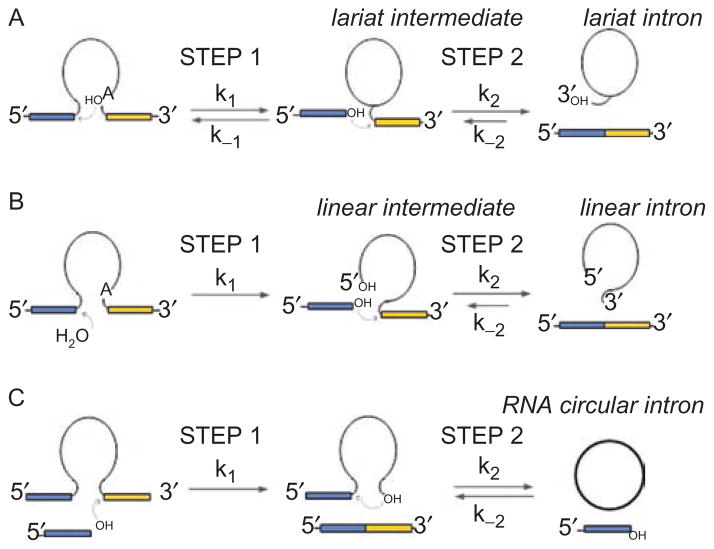
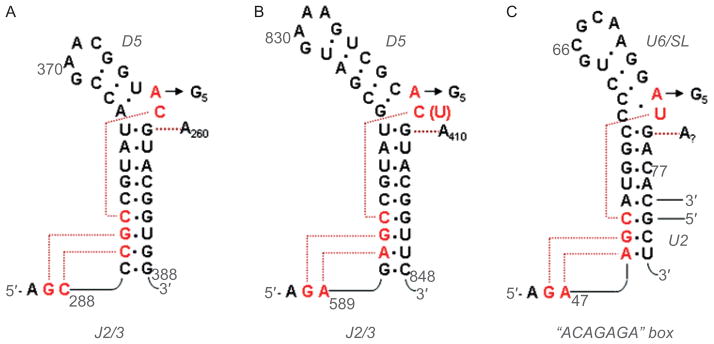
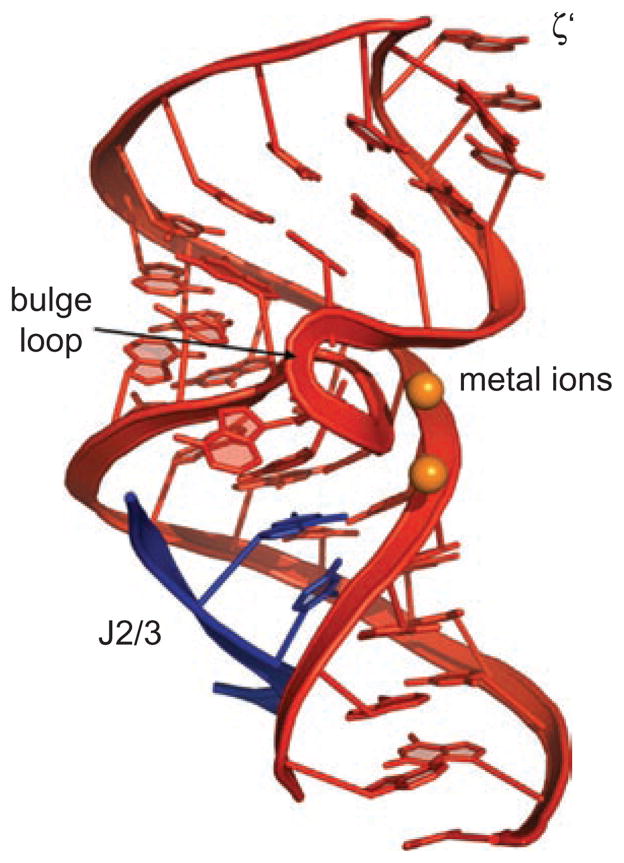
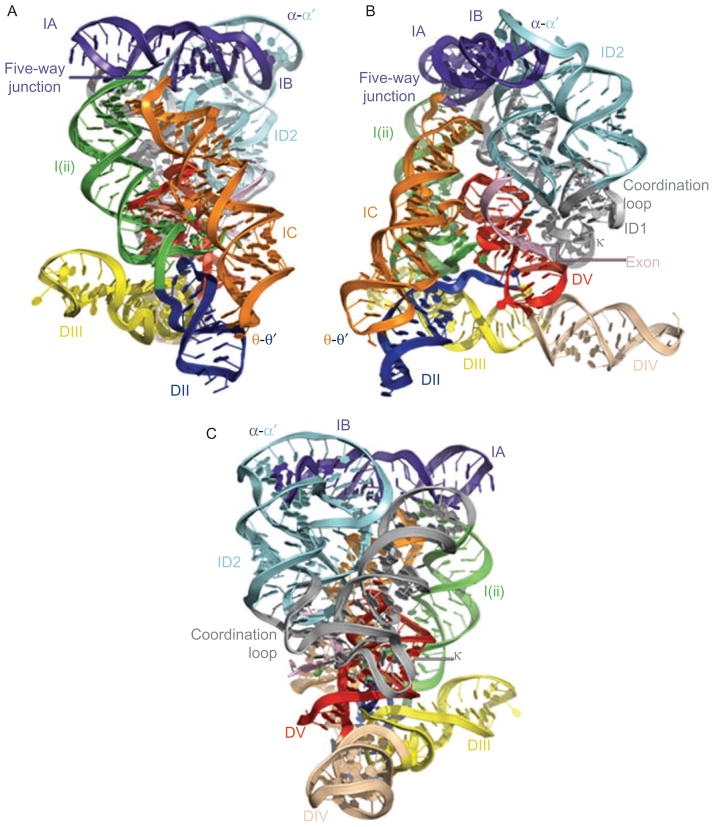
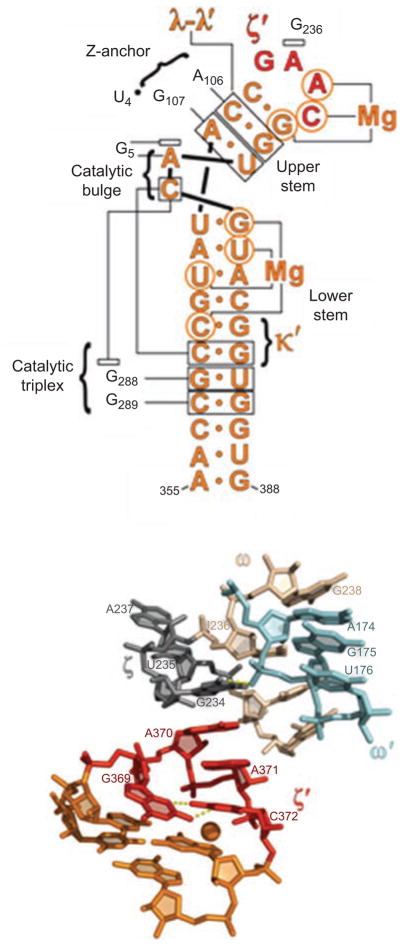
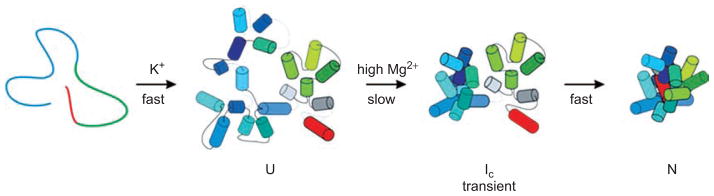
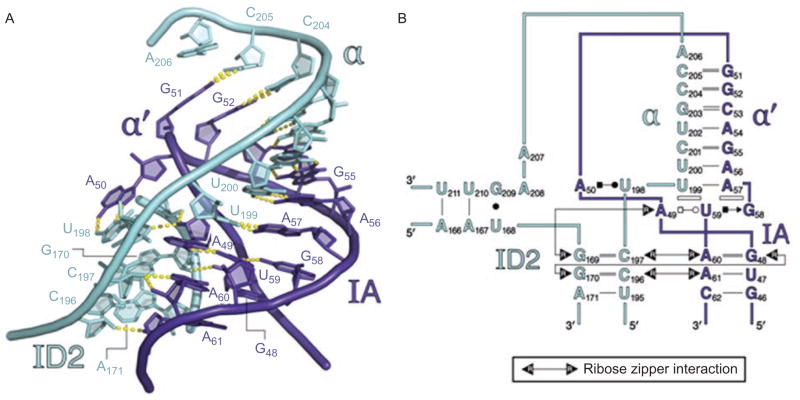
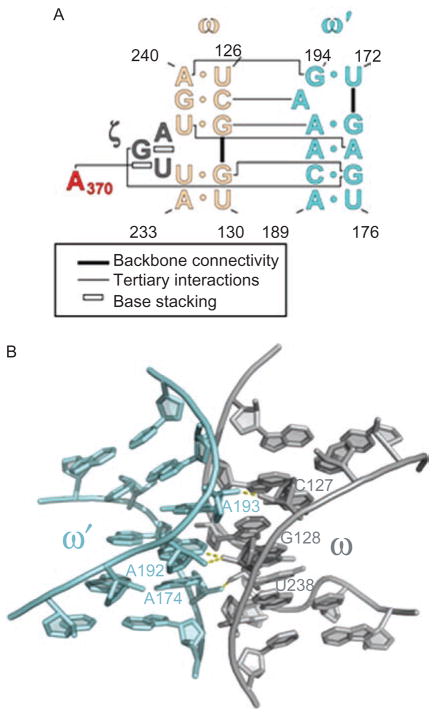
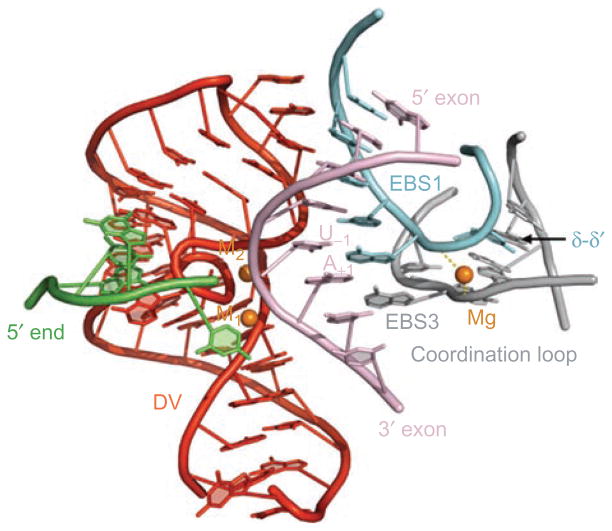
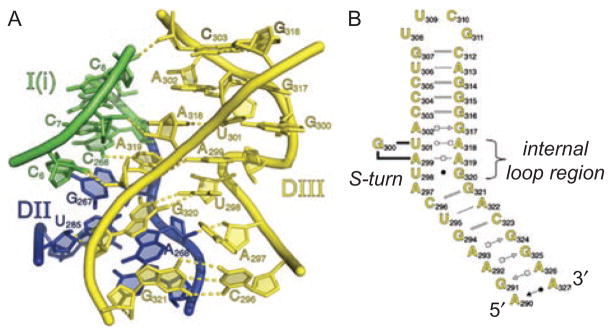
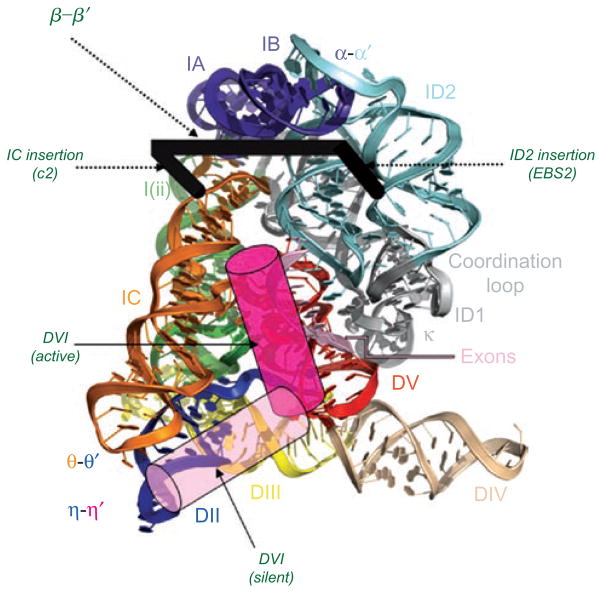
Group II introns are some of the largest ribozymes in nature, and they are a major source of information about RNA assembly and tertiary structural organization. These introns are of biological significance because they are self-splicing mobile elements that have migrated into diverse genomes and played a major role in the genomic organization and metabolism of most life forms. The tertiary structure of group II introns has been the subject of many phylogenetic, genetic, biochemical and biophysical investigations, all of which are consistent with the recent crystal structure of an intact group IIC intron from the alkaliphilic eubacterium Oceanobacillus iheyensis. The crystal structure reveals that catalytic intron domain V is enfolded within the other intronic domains through an elaborate network of diverse tertiary interactions. Within the folded core, DV adopts an activated conformation that readily binds catalytic metal ions and positions them in a manner appropriate for reaction with nucleic acid targets. The tertiary structure of the group II intron reveals new information on motifs for RNA architectural organization, mechanisms of group II intron catalysis, and the evolutionary relationships among RNA processing systems. Guided by the structure and the wealth of previous genetic and biochemical work, it is now possible to deduce the probable location of DVI and the site of additional domains that contribute to the function of the highly derived group IIB and IIA introns.
Conflict of interest statement
I am an Investigator with the Howard Hughes Medical Institute, which supported this work. This work was also supported by a generous grant from the National Institutes of Health (GM50313). The author report no conflicts of interest. The author alone is responsible for the content and writing of the paper.
Figures
References
-
- Abramovitz DL, Friedman RA, Pyle AM. Catalytic role of 2′-hydroxyl groups within a group II intron active site. Science. 1996;271:1410–1413. - PubMed
-
- Adamidi C, Fedorova O, Pyle AM. A group II intron inserted into a bacterial heat-shock operon shows autocatalytic activity and unusual thermostability. Biochemistry. 2003;42:3409–3418. - PubMed
-
- Belfort M, Derbyshire V, Parker M, Cousineau B, Lambowitz A. Mobile introns: pathways and proteins. In: Craig N, Craigie R, Gellert M, Lambowitz A, editors. Mobile DNA II. Washington DC: ASM Press; 2002. pp. 761–783.
-
- Boeke JD. The unusual phylogenetic distribution of retrotransposons: a hypothesis. Genome Res. 2003;13:1975–1983. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources