

Mouse embryonic stem cells, but not somatic cells, predominantly use homologous recombination to repair double-strand DNA breaks
- PMID: 20446816
- PMCID: PMC3128311
- DOI: 10.1089/scd.2010.0058
Mouse embryonic stem cells, but not somatic cells, predominantly use homologous recombination to repair double-strand DNA breaks
Abstract
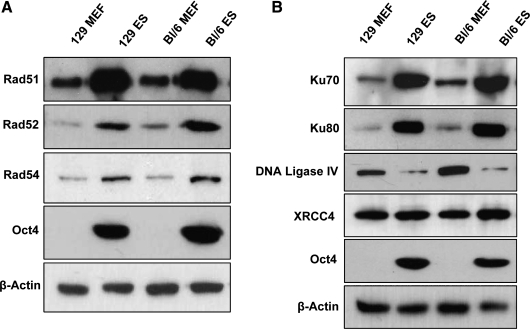
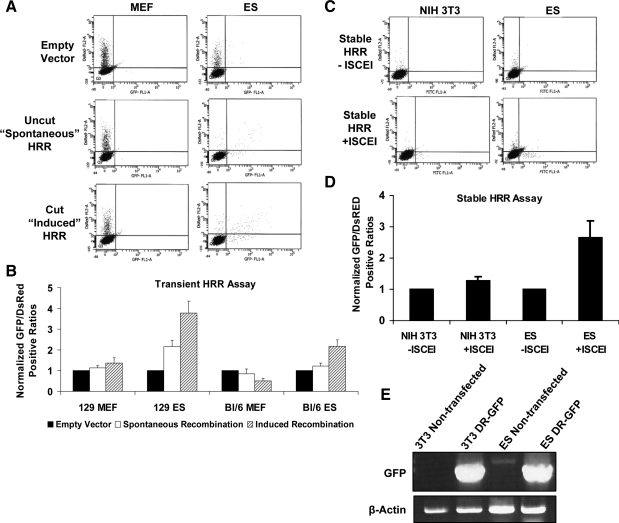
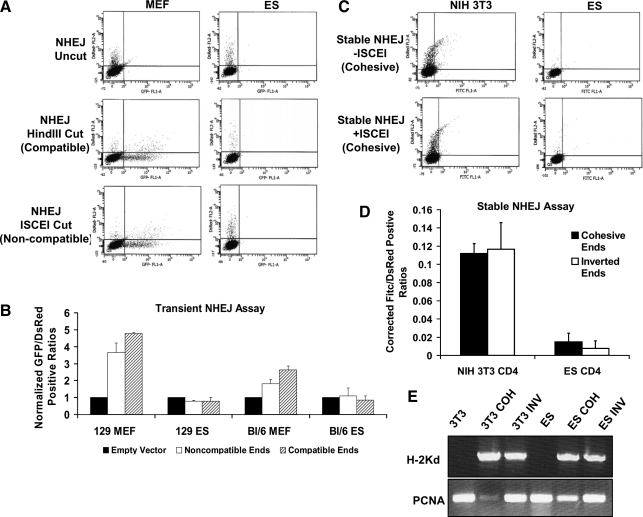
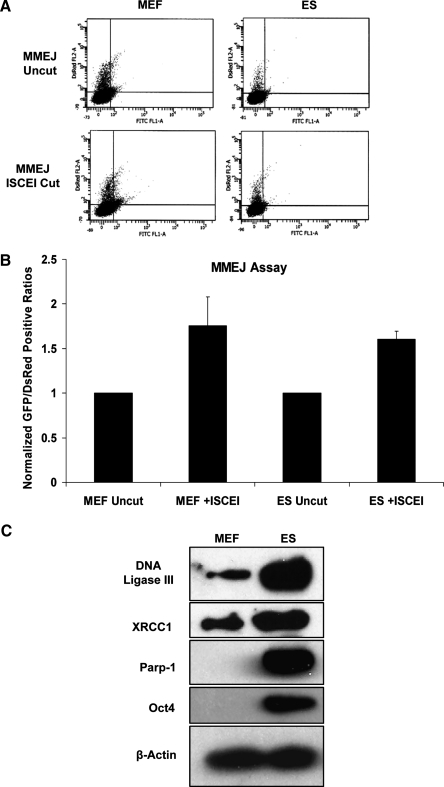
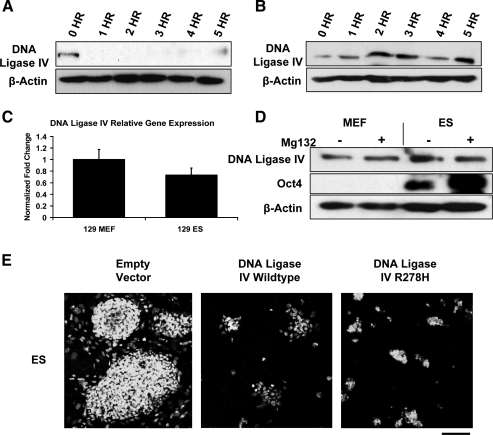
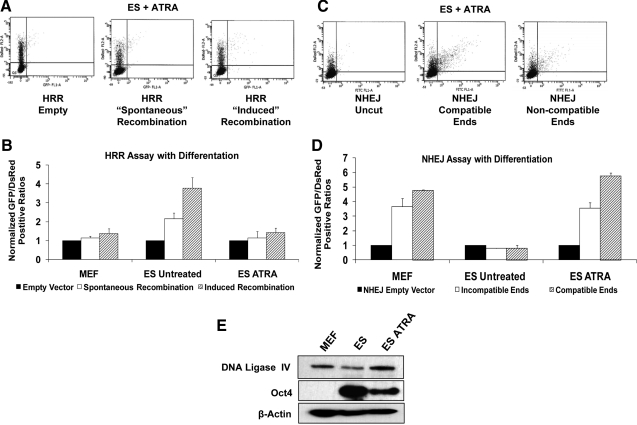
Embryonic stem (ES) cells give rise to all cell types of an organism. Since mutations at this embryonic stage would affect all cells and be detrimental to the overall health of an organism, robust mechanisms must exist to ensure that genomic integrity is maintained. To test this proposition, we compared the capacity of murine ES cells to repair DNA double-strand breaks with that of differentiated cells. Of the 2 major pathways that repair double-strand breaks, error-prone nonhomologous end joining (NHEJ) predominated in mouse embryonic fibroblasts, whereas the high fidelity homologous recombinational repair (HRR) predominated in ES cells. Microhomology-mediated end joining, an emerging repair pathway, persisted at low levels in all cell types examined. The levels of proteins involved in HRR and microhomology-mediated end joining were highly elevated in ES cells compared with mouse embryonic fibroblasts, whereas those for NHEJ were quite variable, with DNA Ligase IV expression low in ES cells. The half-life of DNA Ligase IV protein was also low in ES cells. Attempts to increase the abundance of DNA Ligase IV protein by overexpression or inhibition of its degradation, and thereby elevate NHEJ in ES cells, were unsuccessful. When ES cells were induced to differentiate, however, the level of DNA Ligase IV protein increased, as did the capacity to repair by NHEJ. The data suggest that preferential use of HRR rather than NHEJ may lend ES cells an additional layer of genomic protection and that the limited levels of DNA Ligase IV may account for the low level of NHEJ activity.
Figures
References
-
- Friedberg EC. Meira LB. Database of mouse strains carrying targeted mutations in genes affecting biological responses to DNA damage Version 7. DNA Repair (Amst) 2006;5:189–209. - PubMed
-
- Savatier P. Huang S. Szekely L. Wiman KG. Samarut J. Contrasting patterns of retinoblastoma protein expression in mouse embryonic stem cells and embryonic fibroblasts. Oncogene. 1994;9:809–818. - PubMed
-
- Savatier P. Lapillonne H. Jirmanova L. Vitelli L. Samarut J. Analysis of the cell cycle in mouse embryonic stem cells. Methods Mol Biol. 2002;185:27–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
