

Molecular evolution of piRNA and transposon control pathways in Drosophila
- PMID: 20453205
- PMCID: PMC3181074
- DOI: 10.1101/sqb.2009.74.052
Molecular evolution of piRNA and transposon control pathways in Drosophila
Abstract
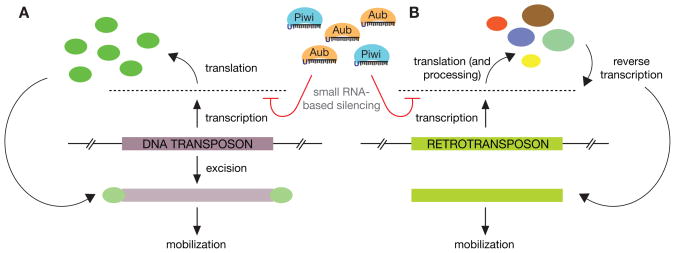
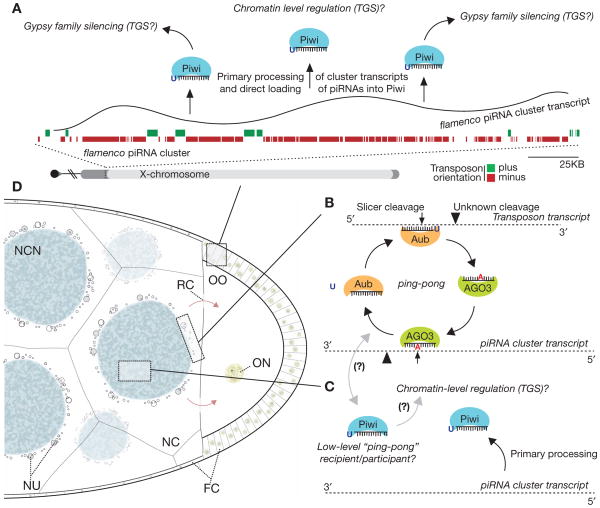
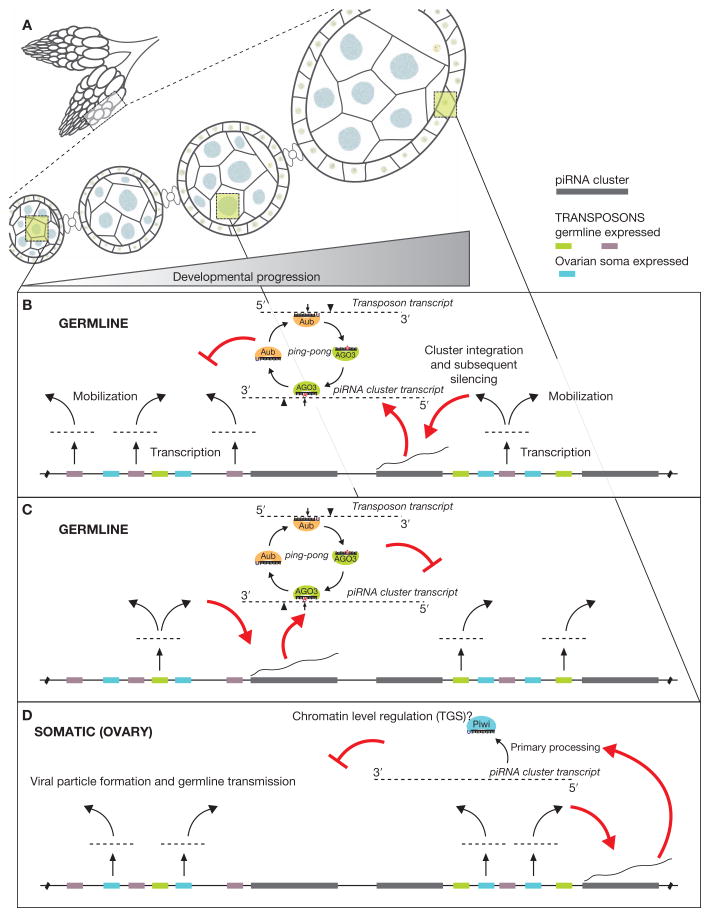
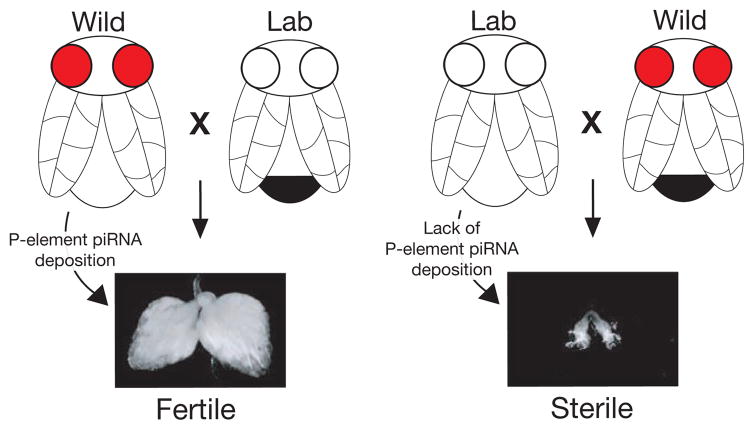
The mere prevalence and potential mobilization of transposable elements in eukaryotic genomes present challenges at both the organismal and population levels. Not only is transposition able to alter gene function and chromosomal structure, but loss of control over even a single active element in the germline can create an evolutionary dead end. Despite the dangers of coexistence, transposons and their activity have been shown to drive the evolution of gene function, chromosomal organization, and even population dynamics (Kazazian 2004). This implies that organisms have adopted elaborate means to balance both the positive and detrimental consequences of transposon activity. In this chapter, we focus on the fruit fly to explore some of the molecular clues into the long- and short-term adaptation to transposon colonization and persistence within eukaryotic genomes.
Figures
References
-
- Aravin AA, Naumova NM, Tulin AV, Vagin VV, Rozovsky YM, Gvozdev VA. Double-stranded RNA-mediated silencing of genomic tandem repeats and transposable elements in the D. melanogaster germline. Curr Biol. 2001;11:1017–1027. - PubMed
-
- Aravin AA, Lagos-Quintana M, Yalcin A, Zavolan M, Marks D, Snyder B, Gaasterland T, Meyer J, Tuschl T. The small RNA profile during Drosophila melanogaster development. Dev Cell. 2003;5:337–350. - PubMed
-
- Aravin A, Gaidatzis D, Pfeffer S, Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ, Kuramochi-Miyagawa S, Nakano T, et al. A novel class of small RNAs bind to MILI protein in mouse testes. Nature. 2006;442:203–207. - PubMed
-
- Aravin AA, Sachidanandam R, Girard A, Fejes-Toth K, Hannon GJ. Developmentally regulated piRNA clusters implicate MILI in transposon control. Science. 2007;316:744–747. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases