

PD-1 regulates germinal center B cell survival and the formation and affinity of long-lived plasma cells
- PMID: 20453843
- PMCID: PMC2874069
- DOI: 10.1038/ni.1877
PD-1 regulates germinal center B cell survival and the formation and affinity of long-lived plasma cells
Abstract
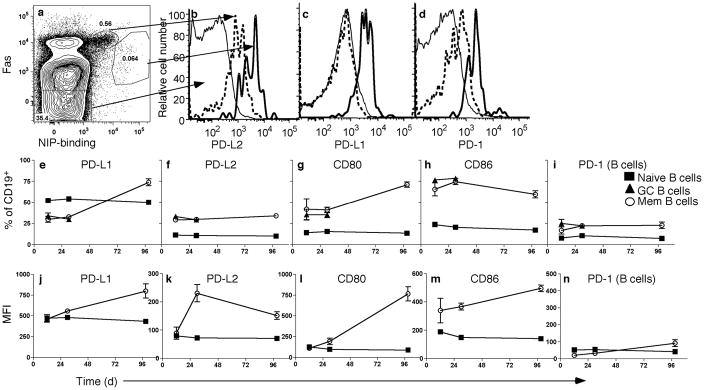
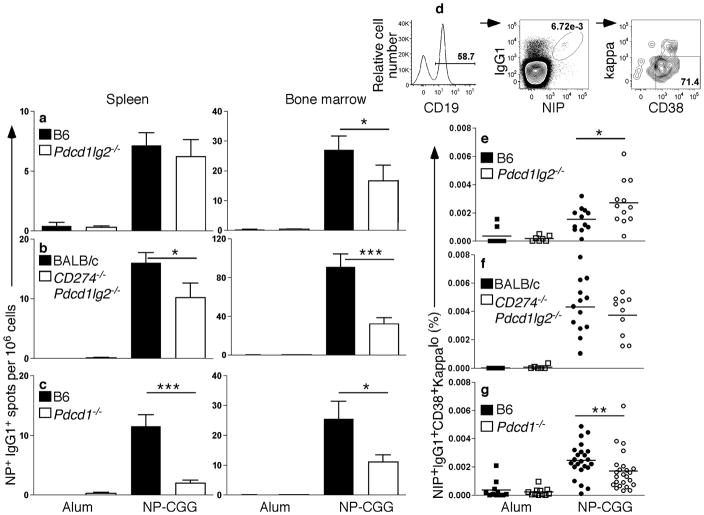
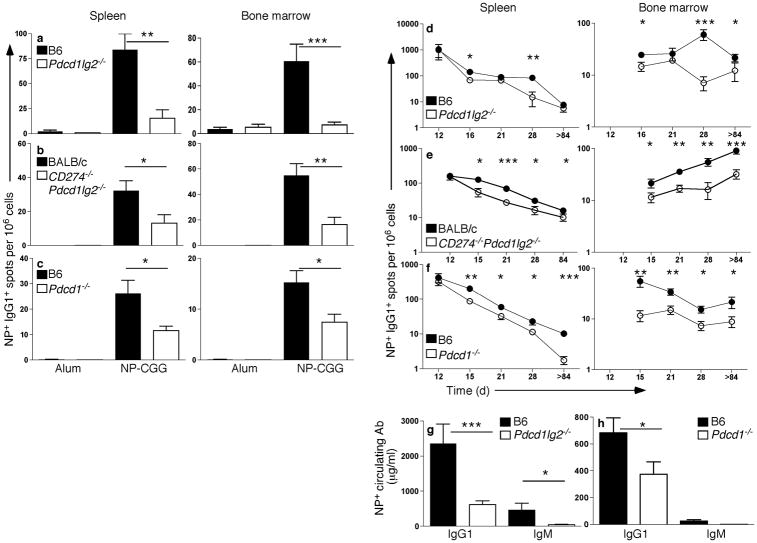
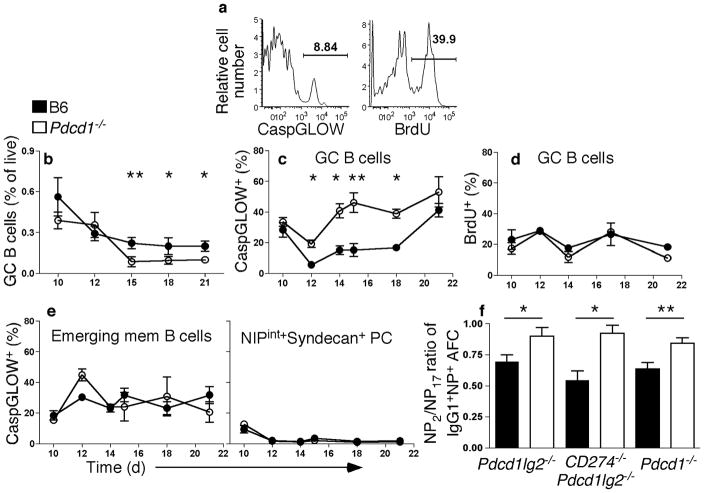
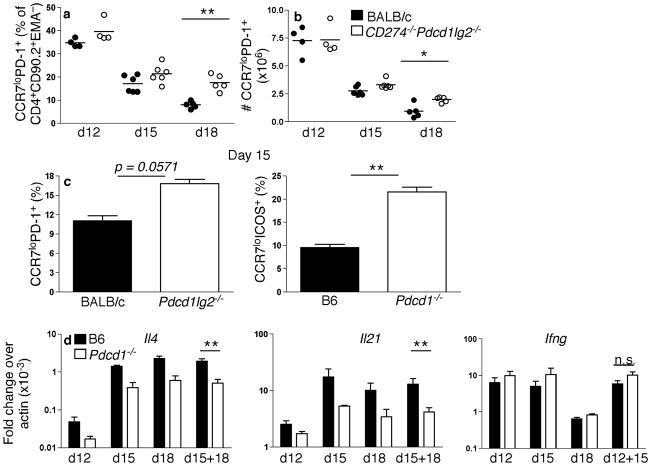
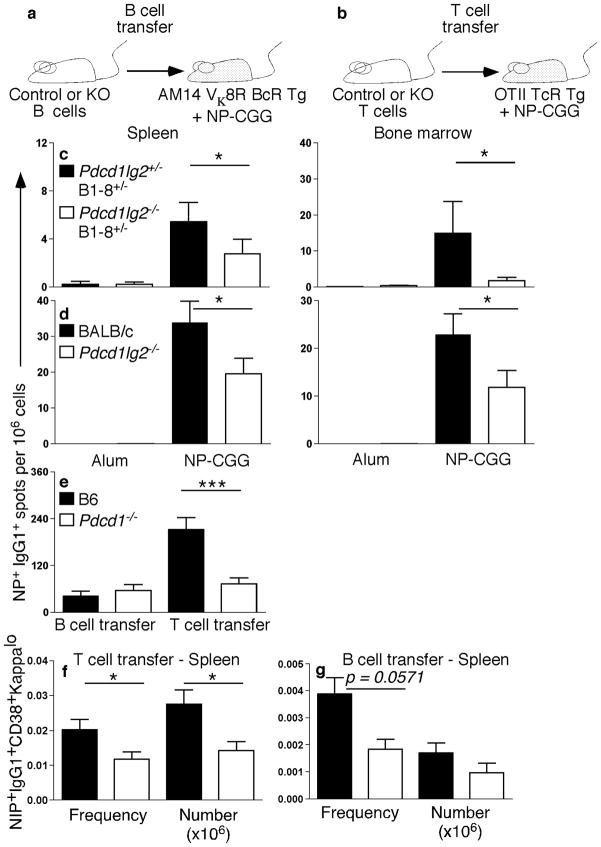
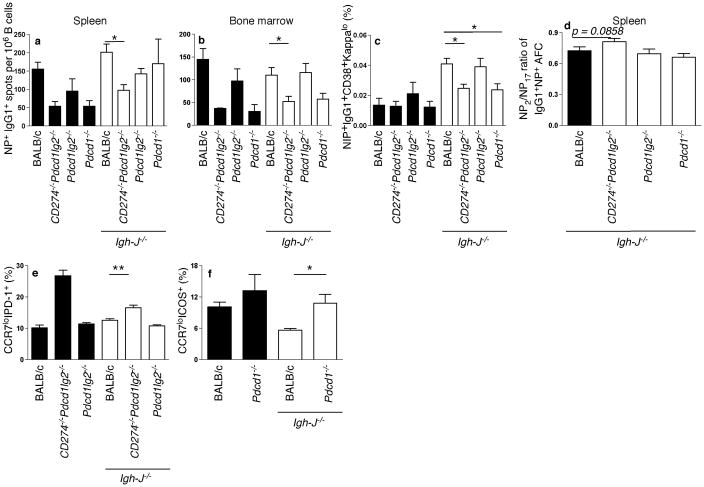
Memory B and plasma cells (PCs) are generated in the germinal center (GC). Because follicular helper T cells (T(FH) cells) have high expression of the immunoinhibitory receptor PD-1, we investigated the role of PD-1 signaling in the humoral response. We found that the PD-1 ligands PD-L1 and PD-L2 were upregulated on GC B cells. Mice deficient in PD-L2 (Pdcd1lg2(-/-)), PD-L1 and PD-L2 (Cd274(-/-)Pdcd1lg2(-/-)) or PD-1 (Pdcd1(-/-)) had fewer long-lived PCs. The mechanism involved more GC cell death and less T(FH) cell cytokine production in the absence of PD-1; the effect was selective, as remaining PCs had greater affinity for antigen. PD-1 expression on T cells and PD-L2 expression on B cells controlled T(FH) cell and PC numbers. Thus, PD-1 regulates selection and survival in the GC, affecting the quantity and quality of long-lived PCs.
Conflict of interest statement
The authors have no conflicts of interest.
Figures
Comment in
-
Give and take in the germinal center.Nat Immunol. 2010 Jun;11(6):464-6. doi: 10.1038/ni0610-464. Nat Immunol. 2010. PMID: 20485272 No abstract available.
References
-
- Joshi NS, Kaech SM. Effector CD8 T cell development: a balancing act between memory cell potential and terminal differentiation. J Immunol. 2008;180:1309–1315. - PubMed
-
- Han S, et al. Cellular interaction in germinal centers. Roles of CD40 ligand and B7-2 in established germinal centers. Journal of Immunology. 1995;155:556–567. - PubMed
-
- Jacob J, Kelsoe G, Rajewsky K, Weiss U. Intraclonal generation of antibody mutants in germinal centres. Nature. 1991;354:389–392. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
