

p53 activates transcription by directing structural shifts in Mediator
- PMID: 20453859
- PMCID: PMC2932482
- DOI: 10.1038/nsmb.1816
p53 activates transcription by directing structural shifts in Mediator
Abstract
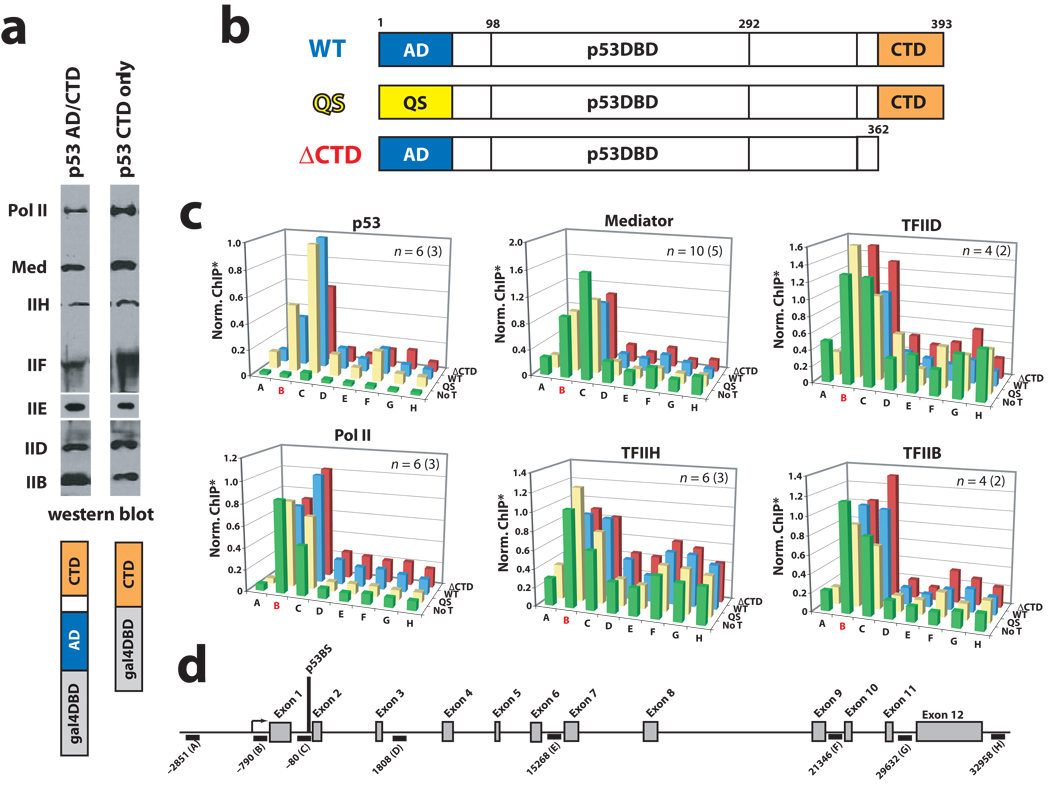
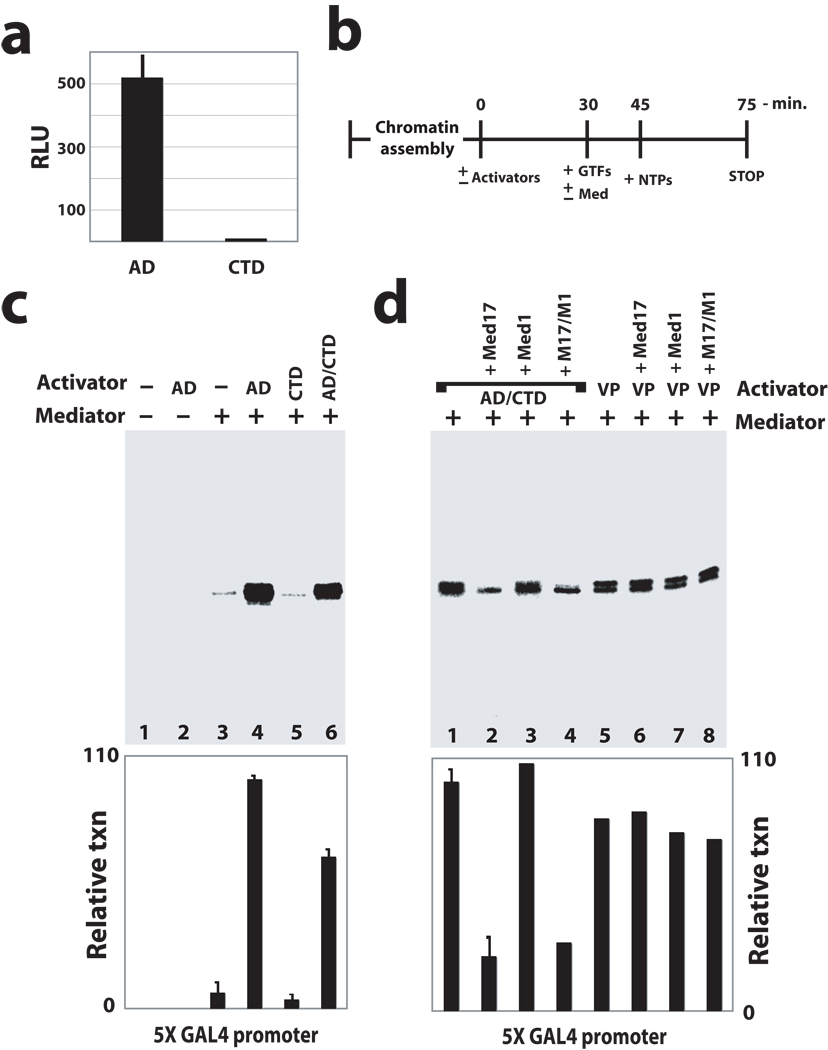
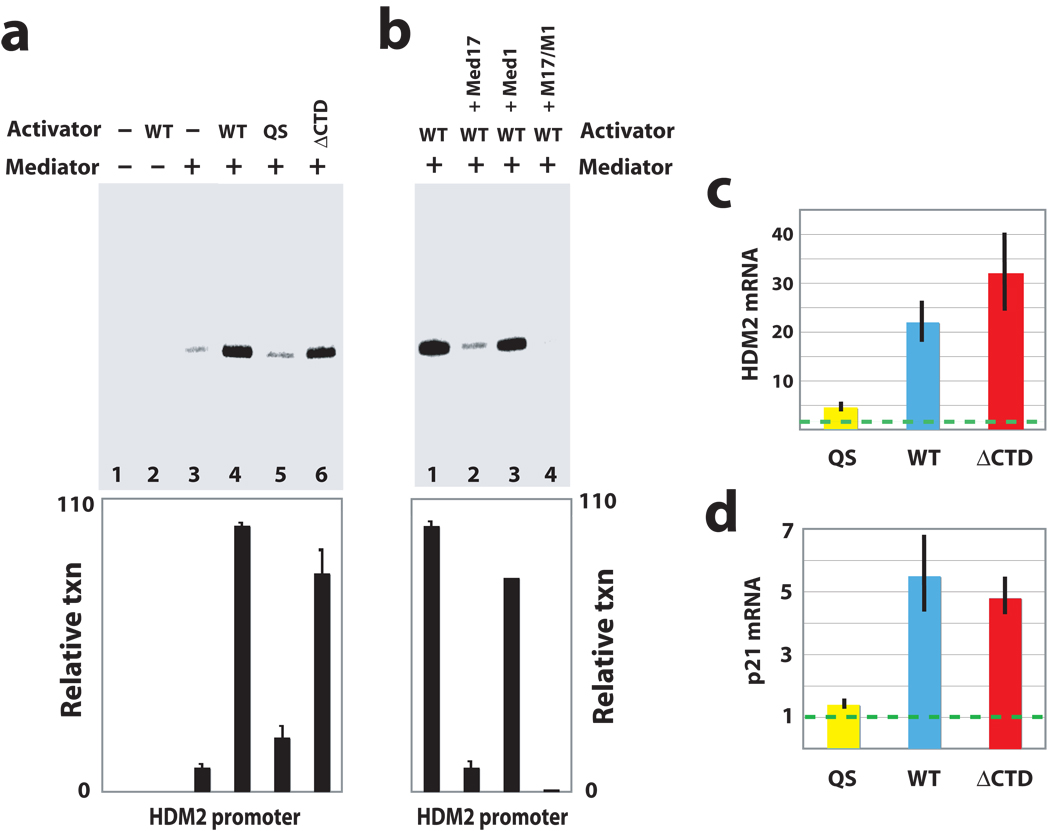
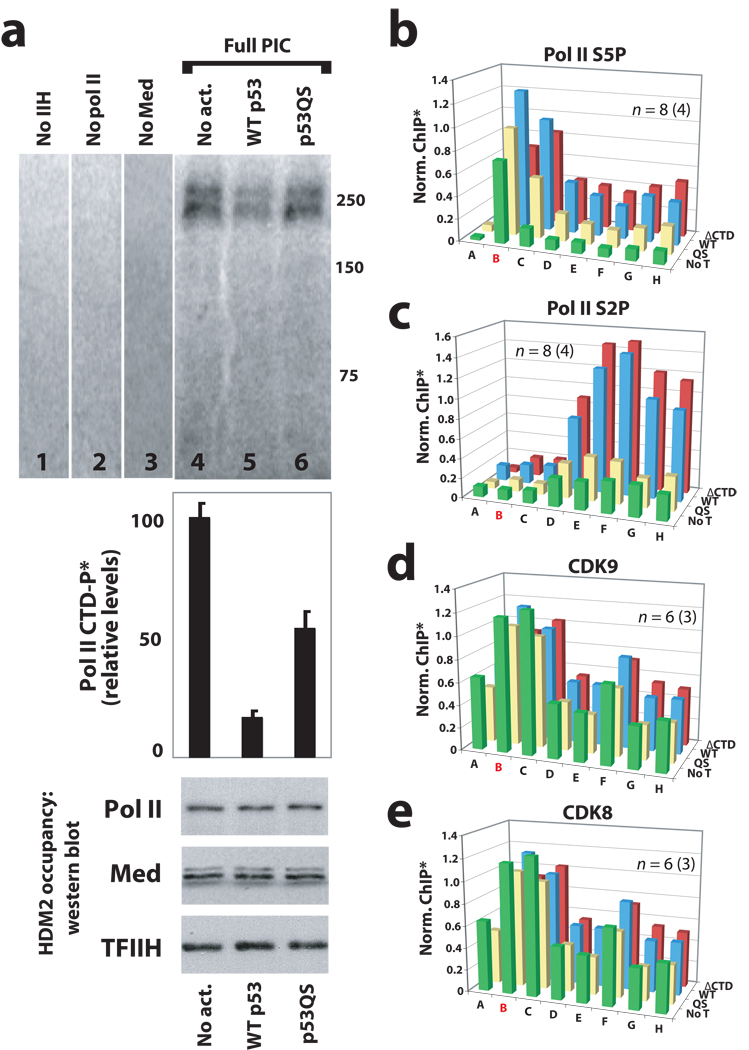
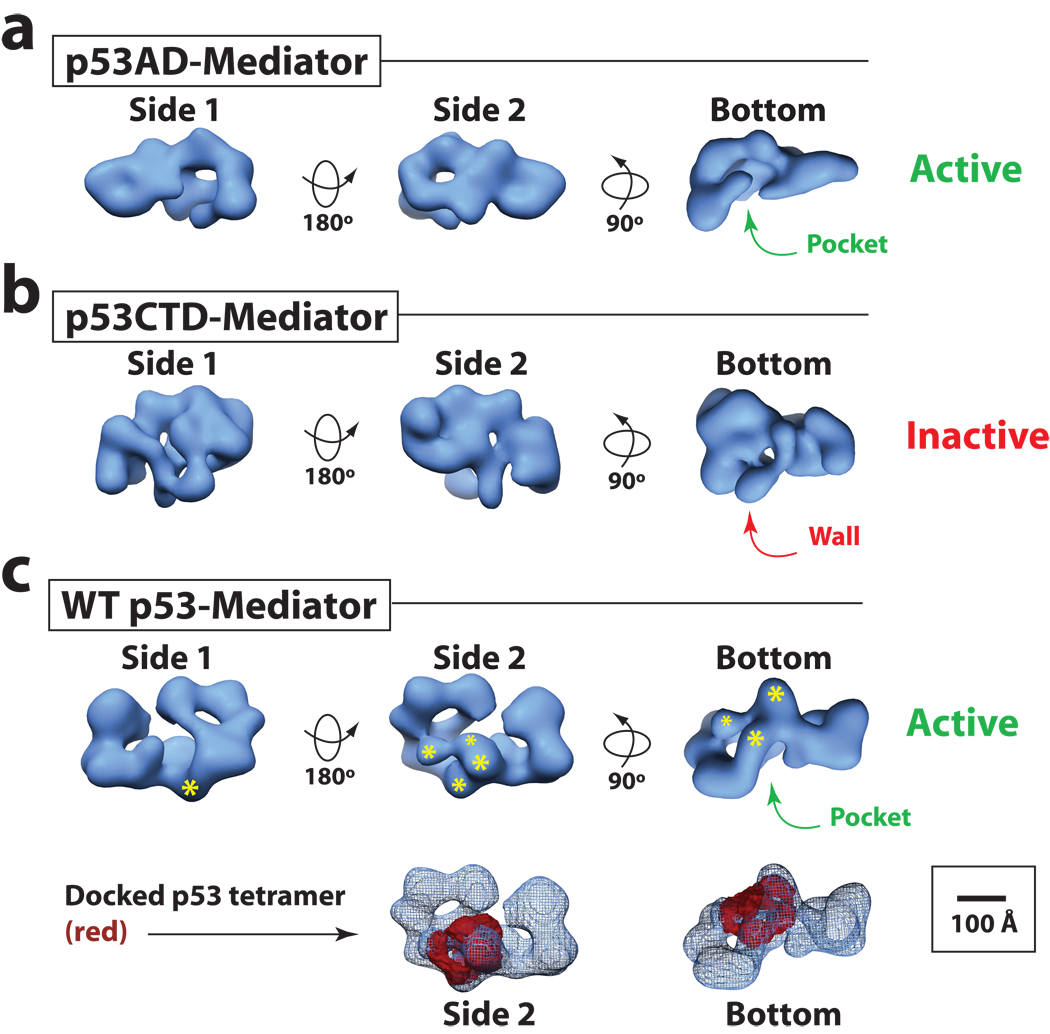
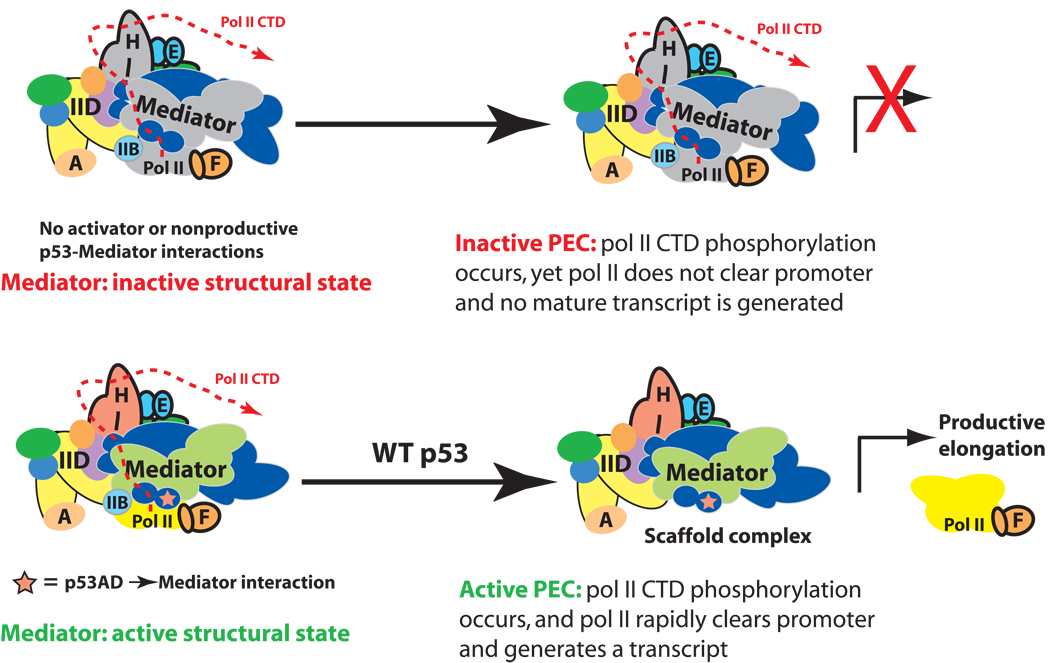
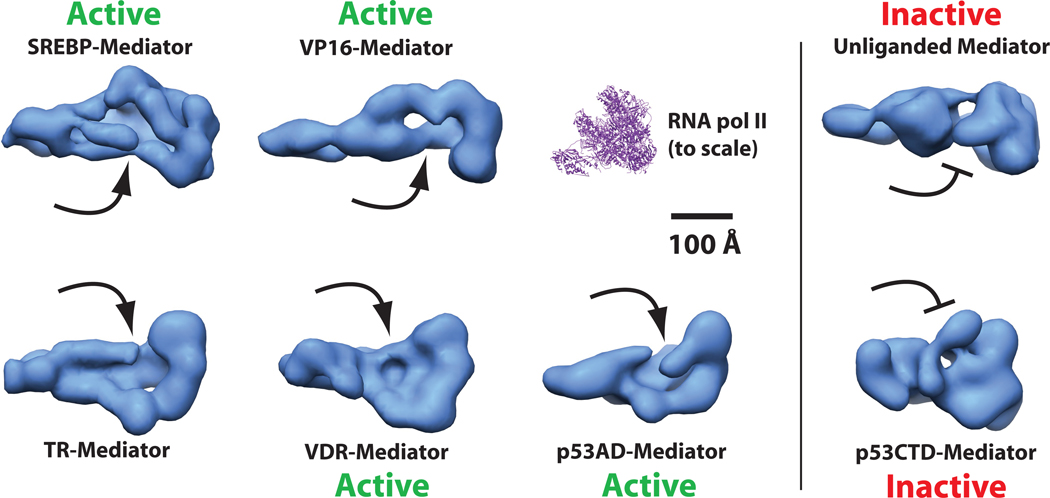
It is not well understood how the human Mediator complex, transcription factor IIH and RNA polymerase II (Pol II) work together with activators to initiate transcription. Activator binding alters Mediator structure, yet the functional consequences of such structural shifts remain unknown. The p53 C terminus and its activation domain interact with different Mediator subunits, and we find that each interaction differentially affects Mediator structure; strikingly, distinct p53-Mediator structures differentially affect Pol II activity. Only the p53 activation domain induces the formation of a large pocket domain at the Mediator-Pol II interaction site, and this correlates with activation of stalled Pol II to a productively elongating state. Moreover, we define a Mediator requirement for TFIIH-dependent Pol II C-terminal domain phosphorylation and identify substantial differences in Pol II C-terminal domain processing that correspond to distinct p53-Mediator structural states. Our results define a fundamental mechanism by which p53 activates transcription and suggest that Mediator structural shifts trigger activation of stalled Pol II complexes.
Figures
Similar articles
-
Origins and activity of the Mediator complex.Semin Cell Dev Biol. 2011 Sep;22(7):729-34. doi: 10.1016/j.semcdb.2011.07.021. Epub 2011 Jul 28. Semin Cell Dev Biol. 2011. PMID: 21821140 Free PMC article. Review.
-
Architecture of the RNA polymerase II-Mediator core initiation complex.Nature. 2015 Feb 19;518(7539):376-80. doi: 10.1038/nature14229. Epub 2015 Feb 4. Nature. 2015. PMID: 25652824
-
Mediator and post-recruitment regulation of RNA polymerase II.Transcription. 2011 Jan-Feb;2(1):28-31. doi: 10.4161/trns.2.1.13950. Transcription. 2011. PMID: 21326907 Free PMC article.
-
Core Mediator structure at 3.4 Å extends model of transcription initiation complex.Nature. 2017 May 11;545(7653):248-251. doi: 10.1038/nature22328. Epub 2017 May 3. Nature. 2017. PMID: 28467824
-
The Mediator complex and transcription elongation.Biochim Biophys Acta. 2013 Jan;1829(1):69-75. doi: 10.1016/j.bbagrm.2012.08.017. Epub 2012 Sep 13. Biochim Biophys Acta. 2013. PMID: 22983086 Free PMC article. Review.
Cited by
-
Mediator complex recruits epigenetic regulators via its two cyclin-dependent kinase subunits to repress transcription of immune response genes.J Biol Chem. 2013 Jul 19;288(29):20955-20965. doi: 10.1074/jbc.M113.486746. Epub 2013 Jun 9. J Biol Chem. 2013. PMID: 23749998 Free PMC article.
-
Mediator head subcomplex Med11/22 contains a common helix bundle building block with a specific function in transcription initiation complex stabilization.Nucleic Acids Res. 2011 Aug;39(14):6291-304. doi: 10.1093/nar/gkr229. Epub 2011 Apr 15. Nucleic Acids Res. 2011. PMID: 21498544 Free PMC article.
-
A combinatorial view of old and new RNA polymerase II modifications.Transcription. 2020 Apr;11(2):66-82. doi: 10.1080/21541264.2020.1762468. Epub 2020 May 13. Transcription. 2020. PMID: 32401151 Free PMC article. Review.
-
The essential and multifunctional TFIIH complex.Protein Sci. 2018 Jun;27(6):1018-1037. doi: 10.1002/pro.3424. Epub 2018 Apr 27. Protein Sci. 2018. PMID: 29664212 Free PMC article. Review.
-
Origins and activity of the Mediator complex.Semin Cell Dev Biol. 2011 Sep;22(7):729-34. doi: 10.1016/j.semcdb.2011.07.021. Epub 2011 Jul 28. Semin Cell Dev Biol. 2011. PMID: 21821140 Free PMC article. Review.
References
-
- Holstege FC, et al. Dissecting the regulatory circuitry of a eukaryotic genome. Cell. 1998;95:717–728. - PubMed
-
- Esnault C, et al. Mediator-dependent recruitment of TFIIH modules in Preinitiation Complex. Mol Cell. 2008;31:337–346. - PubMed
-
- Davis JA, Takagi Y, Kornberg RD, Asturias FA. Structure of the yeast RNA polymerase II holoenzyme: Mediator conformation and polymerase interaction. Mol Cell. 2002;10:409–415. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
