

A novel cancer vaccine strategy based on HLA-A*0201 matched allogeneic plasmacytoid dendritic cells
- PMID: 20454561
- PMCID: PMC2864288
- DOI: 10.1371/journal.pone.0010458
A novel cancer vaccine strategy based on HLA-A*0201 matched allogeneic plasmacytoid dendritic cells
Abstract
Background: The development of effective cancer vaccines still remains a challenge. Despite the crucial role of plasmacytoid dendritic cells (pDCs) in anti-tumor responses, their therapeutic potential has not yet been worked out. We explored the relevance of HLA-A*0201 matched allogeneic pDCs as vectors for immunotherapy.
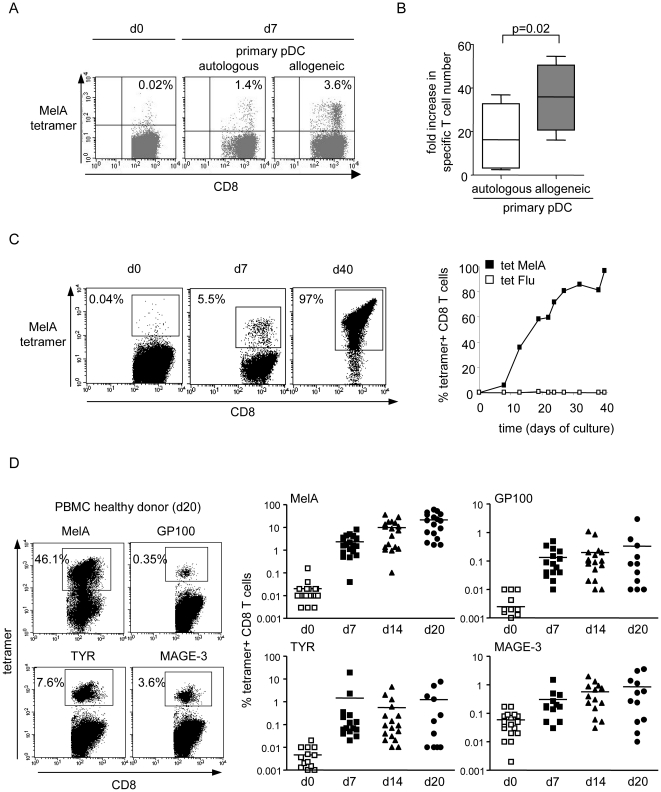
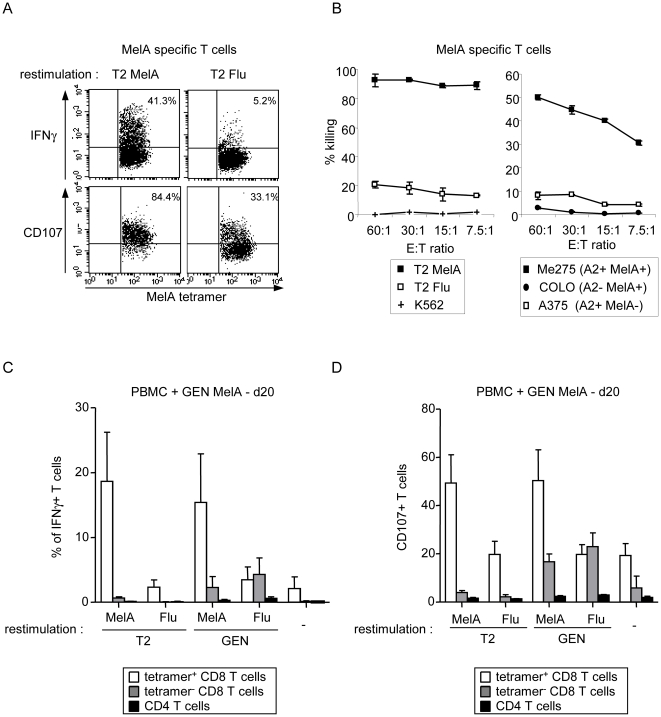
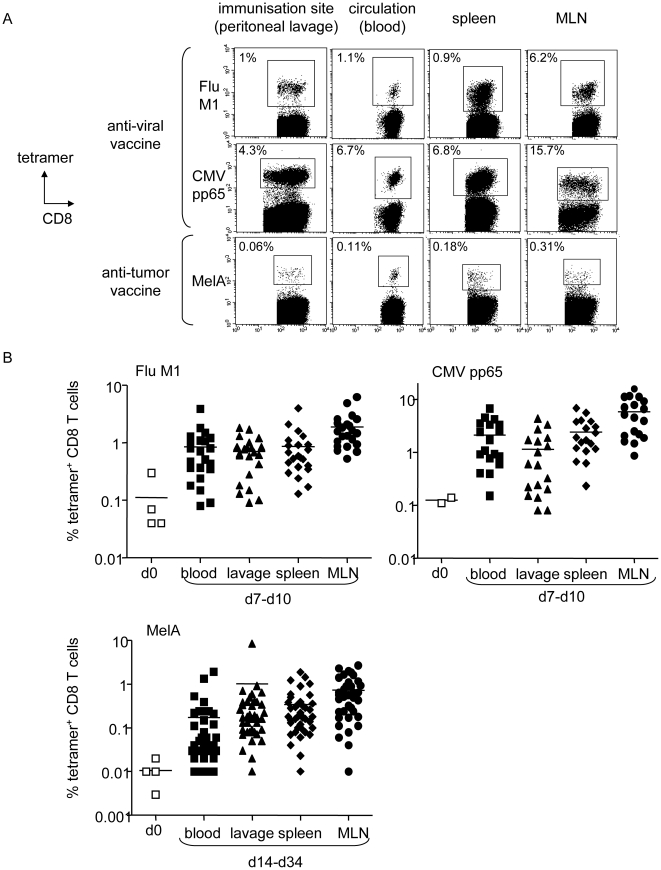
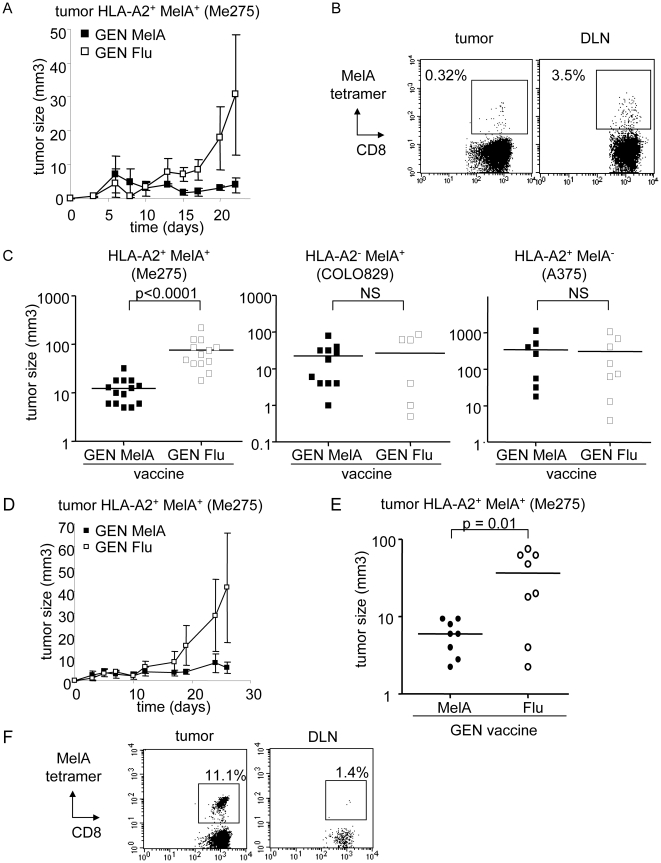
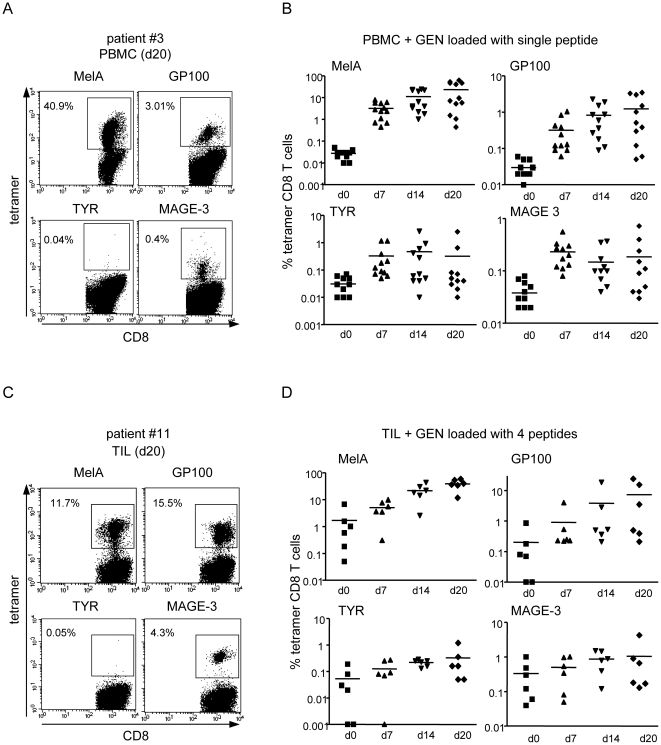
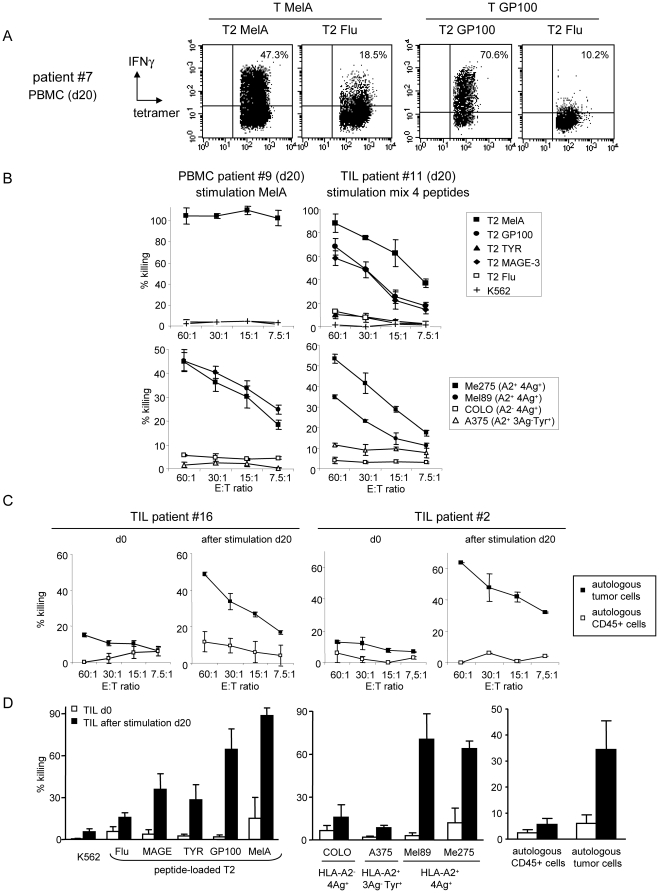
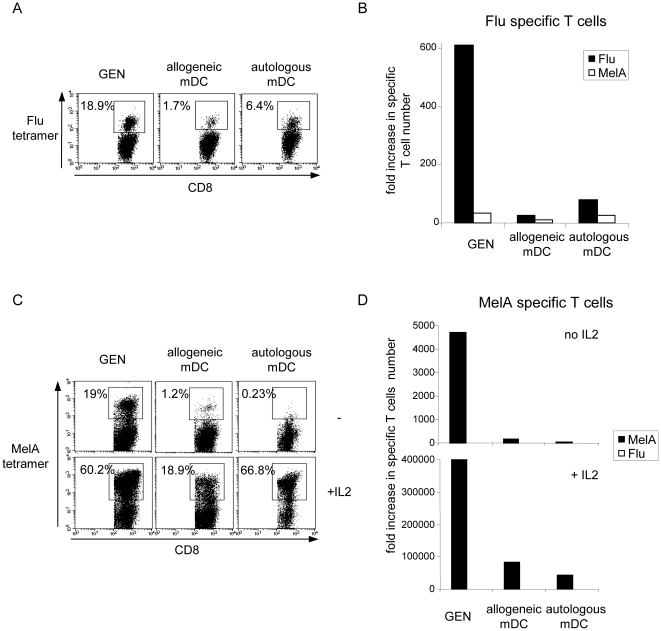
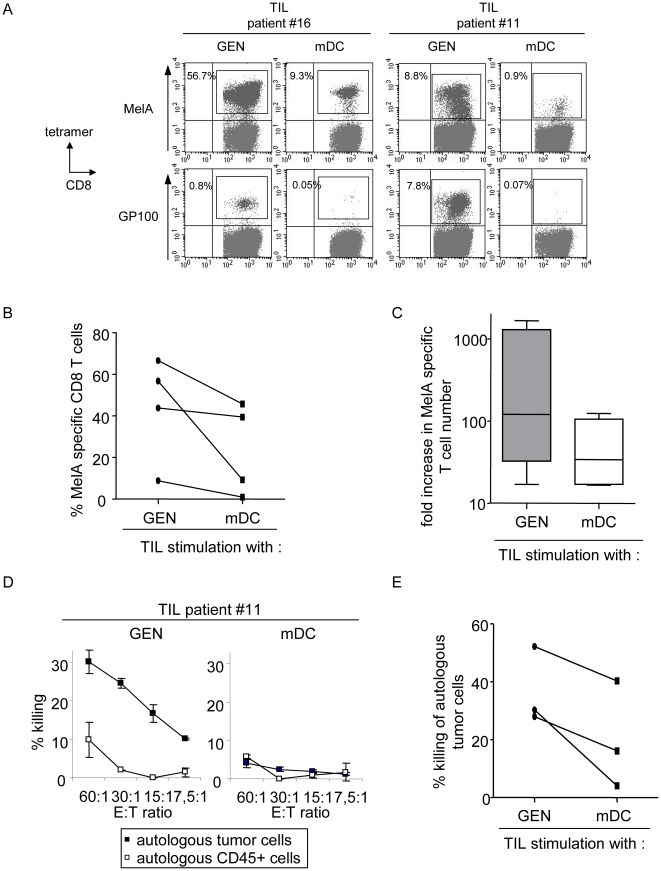
Methods and findings: Stimulation of PBMC from HLA-A*0201(+) donors by HLA-A*0201 matched allogeneic pDCs pulsed with tumor-derived peptides triggered high levels of antigen-specific and functional cytotoxic T cell responses (up to 98% tetramer(+) CD8 T cells). The pDC vaccine demonstrated strong anti-tumor therapeutic in vivo efficacy as shown by the inhibition of tumor growth in a humanized mouse model. It also elicited highly functional tumor-specific T cells ex-vivo from PBMC and TIL of stage I-IV melanoma patients. Responses against MelA, GP100, tyrosinase and MAGE-3 antigens reached tetramer levels up to 62%, 24%, 85% and 4.3% respectively. pDC vaccine-primed T cells specifically killed patients' own autologous melanoma tumor cells. This semi-allogeneic pDC vaccine was more effective than conventional myeloid DC-based vaccines. Furthermore, the pDC vaccine design endows it with a strong potential for clinical application in cancer treatment.
Conclusions: These findings highlight HLA-A*0201 matched allogeneic pDCs as potent inducers of tumor immunity and provide a promising immunotherapeutic strategy to fight cancer.
Conflict of interest statement
Figures
References
-
- Finn OJ. Cancer immunology. N Engl J Med. 2008;358:2704–2715. Review. - PubMed
-
- Thurner B, Haendle I, Röder C, Dieckmann D, Keikavoussi P, et al. Vaccination with mage-3A1 peptide-pulsed mature, monocyte-derived dendritic cells expands specific cytotoxic T cells and induces regression of some metastases in advanced stage IV melanoma. J Exp Med. 1999;190:1669–1678. - PMC - PubMed
-
- Palucka AK, Dhodapkar MV, Paczesny S, Ueno H, Fay J, et al. Boosting vaccinations with peptide-pulsed CD34+ progenitor-derived dendritic cells can expand long-lived melanoma peptide-specific CD8+ T cells in patients with metastatic melanoma. J Immunother. 2005;28:158–168. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
