

A differential role for neuropeptides in acute and chronic adaptive responses to alcohol: behavioural and genetic analysis in Caenorhabditis elegans
- PMID: 20454655
- PMCID: PMC2862703
- DOI: 10.1371/journal.pone.0010422
A differential role for neuropeptides in acute and chronic adaptive responses to alcohol: behavioural and genetic analysis in Caenorhabditis elegans
Abstract
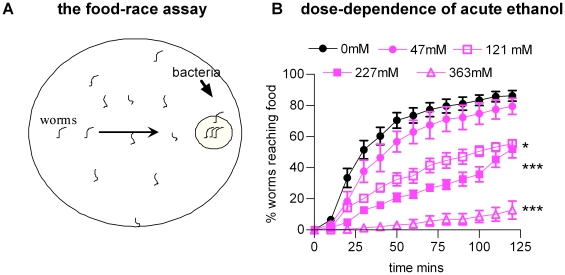
Prolonged alcohol consumption in humans followed by abstinence precipitates a withdrawal syndrome consisting of anxiety, agitation and in severe cases, seizures. Withdrawal is relieved by a low dose of alcohol, a negative reinforcement that contributes to alcohol dependency. This phenomenon of 'withdrawal relief' provides evidence of an ethanol-induced adaptation which resets the balance of signalling in neural circuits. We have used this as a criterion to distinguish between direct and indirect ethanol-induced adaptive behavioural responses in C. elegans with the goal of investigating the genetic basis of ethanol-induced neural plasticity. The paradigm employs a 'food race assay' which tests sensorimotor performance of animals acutely and chronically treated with ethanol. We describe a multifaceted C. elegans 'withdrawal syndrome'. One feature, decrease reversal frequency is not relieved by a low dose of ethanol and most likely results from an indirect adaptation to ethanol caused by inhibition of feeding and a food-deprived behavioural state. However another aspect, an aberrant behaviour consisting of spontaneous deep body bends, did show withdrawal relief and therefore we suggest this is the expression of ethanol-induced plasticity. The potassium channel, slo-1, which is a candidate ethanol effector in C. elegans, is not required for the responses described here. However a mutant deficient in neuropeptides, egl-3, is resistant to withdrawal (although it still exhibits acute responses to ethanol). This dependence on neuropeptides does not involve the NPY-like receptor npr-1, previously implicated in C. elegans ethanol withdrawal. Therefore other neuropeptide pathways mediate this effect. These data resonate with mammalian studies which report involvement of a number of neuropeptides in chronic responses to alcohol including corticotrophin-releasing-factor (CRF), opioids, tachykinins as well as NPY. This suggests an evolutionarily conserved role for neuropeptides in ethanol-induced plasticity and opens the way for a genetic analysis of the effects of alcohol on a simple model system.
Conflict of interest statement
Figures












References
-
- DSM-IV. Diagnostic and Statistical Manual of Mental Disorders of the American Psychiatric Association. 1994. Washington DC.
-
- Weiss F. Neurocircuitry targets in ethanol reward and dependence. Alcohol Clin Exp Res. 1998;22:3–9. - PubMed
-
- Heinz A, Schäfer M, Higley JD, Krystal JH, Goldman D. Neurobiological correlates of the disposition and maintenance of alcoholism. Pharmacopsychiatry. 2003;Suppl 3:S255–S258. - PubMed
-
- Koob GF, Le Moal M. London: Elsevier Inc; 2006. Alcohol. pp. 173–241. In: Neurobiology of Addiction, Koob GF, Le Moal M.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
