

Proteomic profiling of human keratinocytes undergoing UVB-induced alternative differentiation reveals TRIpartite Motif Protein 29 as a survival factor
- PMID: 20454669
- PMCID: PMC2862717
- DOI: 10.1371/journal.pone.0010462
Proteomic profiling of human keratinocytes undergoing UVB-induced alternative differentiation reveals TRIpartite Motif Protein 29 as a survival factor
Abstract
Background: Repeated exposures to UVB of human keratinocytes lacking functional p16(INK-4a) and able to differentiate induce an alternative state of differentiation rather than stress-induced premature senescence.
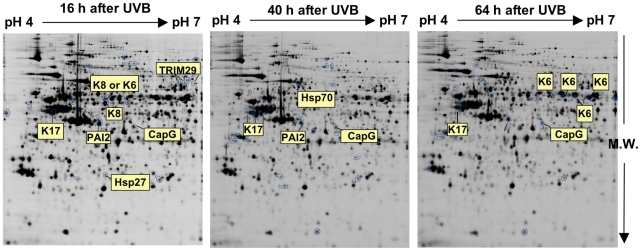
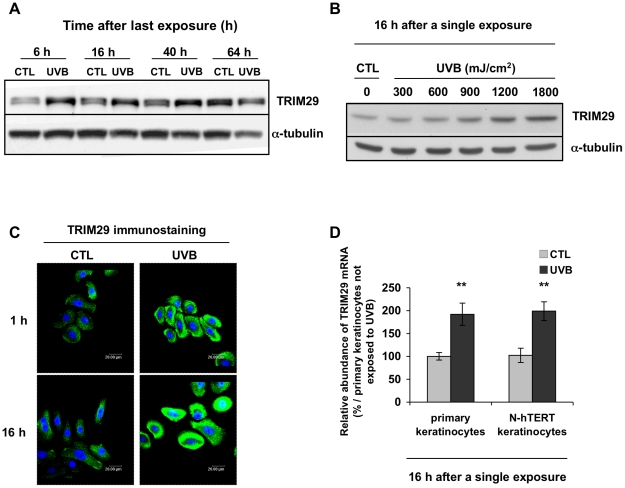
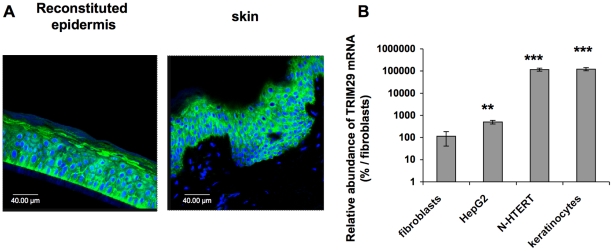
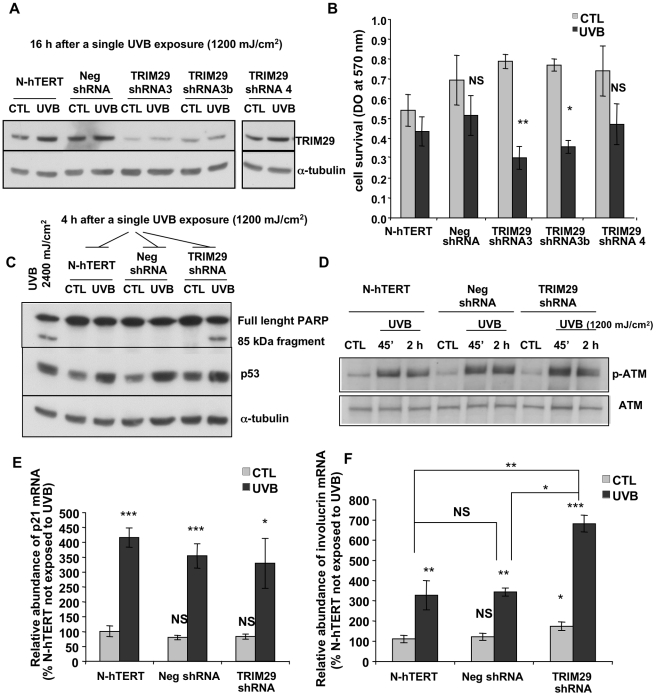
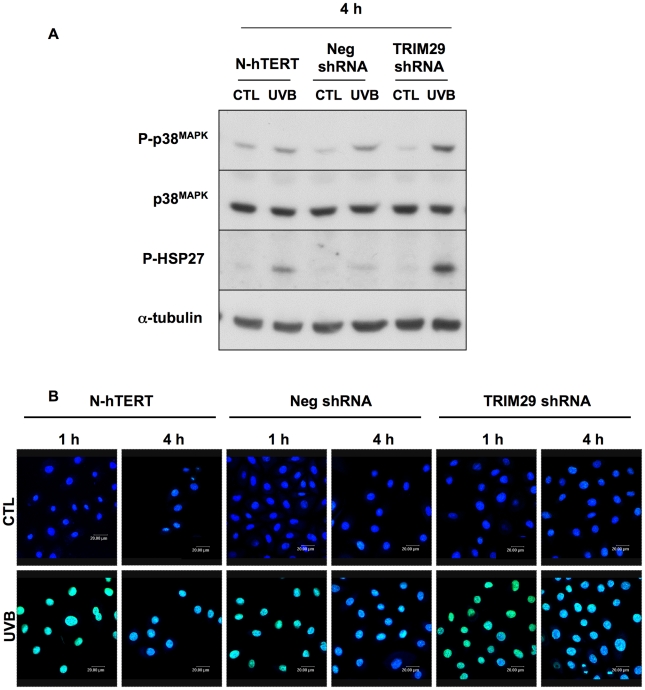
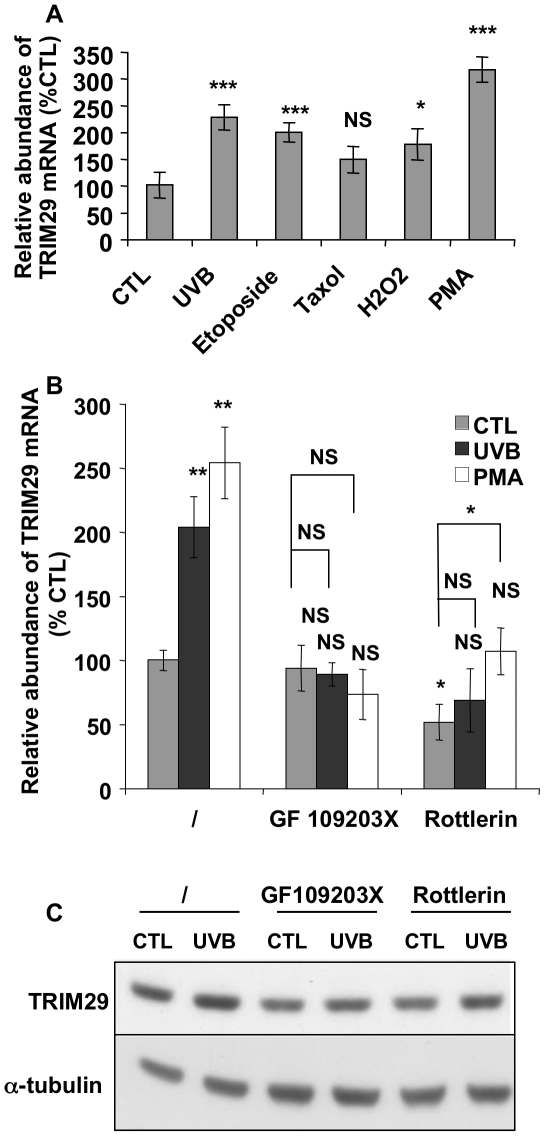
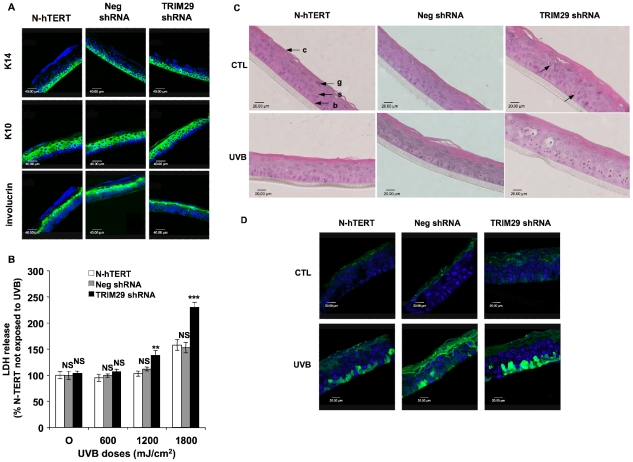
Methodology/principal findings: A 2D-DIGE proteomic profiling of this alternative state of differentiation was performed herein at various times after the exposures to UVB. Sixty-nine differentially abundant protein species were identified by mass spectrometry, many of which are involved in keratinocyte differentiation and survival. Among these protein species was TRIpartite Motif Protein 29 (TRIM29). Increased abundance of TRIM29 following UVB exposures was validated by Western blot using specific antibody and was also further analysed by immunochemistry and by RT-PCR. TRIM29 was found very abundant in keratinocytes and reconstructed epidermis. Knocking down the expression of TRIM29 by short-hairpin RNA interference decreased the viability of keratinocytes after UVB exposure. The abundance of involucrin mRNA, a marker of late differentiation, increased concomitantly. In TRIM29-knocked down reconstructed epidermis, the presence of picnotic cells revealed cell injury. Increased abundance of TRIM29 was also observed upon exposure to DNA damaging agents and PKC activation. The UVB-induced increase of TRIM29 abundance was dependent on a PKC signaling pathway, likely PKCdelta.
Conclusions/significance: These findings suggest that TRIM29 allows keratinocytes to enter a protective alternative differentiation process rather than die massively after stress.
Conflict of interest statement
Figures
Similar articles
-
Repeated exposures to UVB induce differentiation rather than senescence of human keratinocytes lacking p16(INK-4A).Biogerontology. 2010 Apr;11(2):167-81. doi: 10.1007/s10522-009-9238-y. Epub 2009 Jun 24. Biogerontology. 2010. PMID: 19554468
-
Ultraviolet radiation B induces differentiation and protein kinase C in normal human epidermal keratinocytes.Photodermatol Photoimmunol Photomed. 1996 Jun;12(3):103-8. doi: 10.1111/j.1600-0781.1996.tb00185.x. Photodermatol Photoimmunol Photomed. 1996. PMID: 8956359
-
Id2 protein is selectively upregulated by UVB in primary, but not in immortalized human keratinocytes and inhibits differentiation.Oncogene. 2005 Aug 18;24(35):5443-58. doi: 10.1038/sj.onc.1208709. Oncogene. 2005. PMID: 16007217
-
TRIM-endous functional network of tripartite motif 29 (TRIM29) in cancer progression and beyond.Cancer Metastasis Rev. 2024 Dec 7;44(1):16. doi: 10.1007/s10555-024-10226-2. Cancer Metastasis Rev. 2024. PMID: 39644332 Free PMC article. Review.
-
TRIM29 in Cutaneous Squamous Cell Carcinoma.Front Med (Lausanne). 2021 Dec 20;8:804166. doi: 10.3389/fmed.2021.804166. eCollection 2021. Front Med (Lausanne). 2021. PMID: 34988104 Free PMC article. Review.
Cited by
-
ATDC (Ataxia Telangiectasia Group D Complementing) Promotes Radioresistance through an Interaction with the RNF8 Ubiquitin Ligase.J Biol Chem. 2015 Nov 6;290(45):27146-27157. doi: 10.1074/jbc.M115.665489. Epub 2015 Sep 17. J Biol Chem. 2015. PMID: 26381412 Free PMC article.
-
Immortalized N/TERT keratinocytes as an alternative cell source in 3D human epidermal models.Sci Rep. 2017 Sep 19;7(1):11838. doi: 10.1038/s41598-017-12041-y. Sci Rep. 2017. PMID: 28928444 Free PMC article.
-
Silencing of tripartite motif (TRIM) 29 inhibits proliferation and invasion and increases chemosensitivity to cisplatin in human lung squamous cancer NCI-H520 cells.Thorac Cancer. 2015 Jan;6(1):31-7. doi: 10.1111/1759-7714.12130. Epub 2015 Jan 7. Thorac Cancer. 2015. PMID: 26273332 Free PMC article.
-
Terminal keratinocyte differentiation in vitro is associated with a stable DNA methylome.Exp Dermatol. 2021 Aug;30(8):1023-1032. doi: 10.1111/exd.14153. Epub 2020 Jul 28. Exp Dermatol. 2021. PMID: 32681572 Free PMC article.
-
UV, stress and aging.Dermatoendocrinol. 2012 Jul 1;4(3):236-40. doi: 10.4161/derm.23652. Dermatoendocrinol. 2012. PMID: 23467762 Free PMC article.
References
-
- Eckert RL, Crish JF, Robinson NA. The epidermal keratinocyte as a model for the study of gene regulation and cell differentiation. Physiol Rev. 1997;77:397–424. - PubMed
-
- Ichihashi M, Ueda M, Budiyanto A, Bito T, Oka M, et al. UV-induced skin damage. Toxicology. 2003;189:21–39. - PubMed
-
- Svobodova A, Walterova D, Vostalova J. Ultraviolet light induced alteration to the skin. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2006;150:25–38. - PubMed
-
- Rittie L, Fisher GJ. UV-light-induced signal cascades and skin aging. Ageing Res Rev. 2002;1:705–720. - PubMed
-
- D'Errico M, Teson M, Calcagnile A, Proietti De Santis L, Nikaido O, et al. Apoptosis and efficient repair of DNA damage protect human keratinocytes against UVB. Cell Death Differ. 2003;10:754–756. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
