

A novel single-stranded DNA-specific 3'-5' exonuclease, Thermus thermophilus exonuclease I, is involved in several DNA repair pathways
- PMID: 20457749
- PMCID: PMC2943613
- DOI: 10.1093/nar/gkq350
A novel single-stranded DNA-specific 3'-5' exonuclease, Thermus thermophilus exonuclease I, is involved in several DNA repair pathways
Abstract
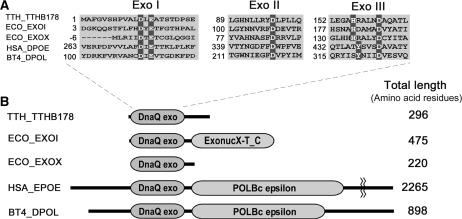
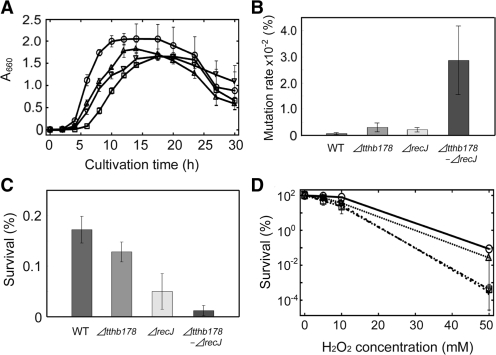
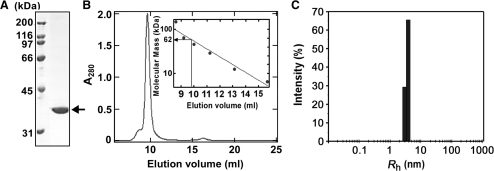
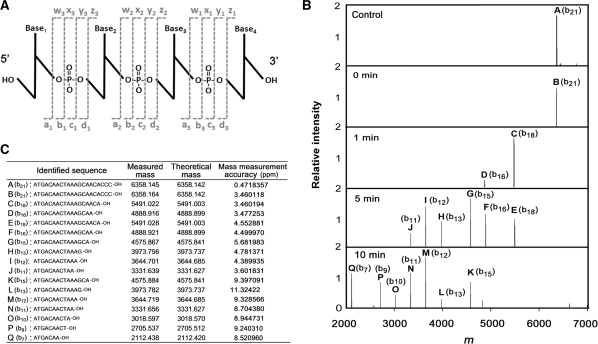
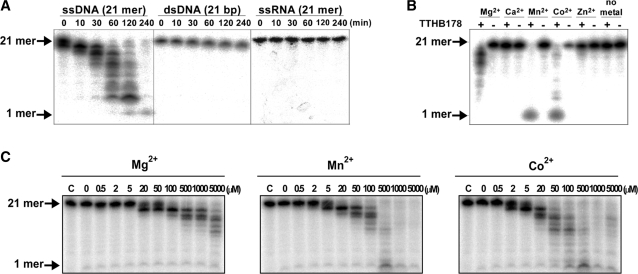
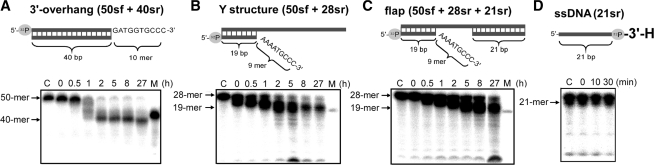
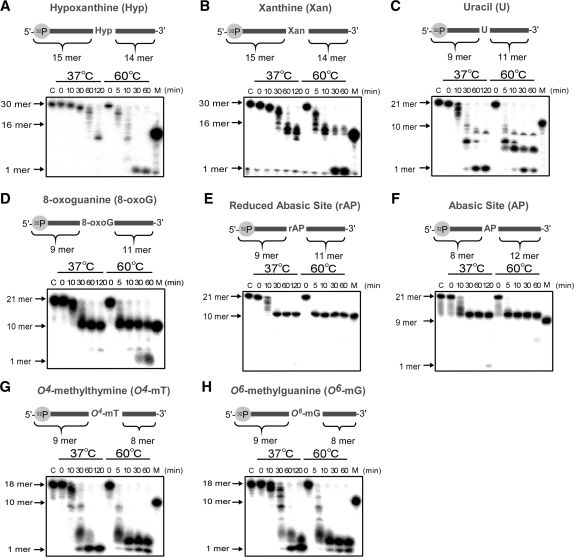
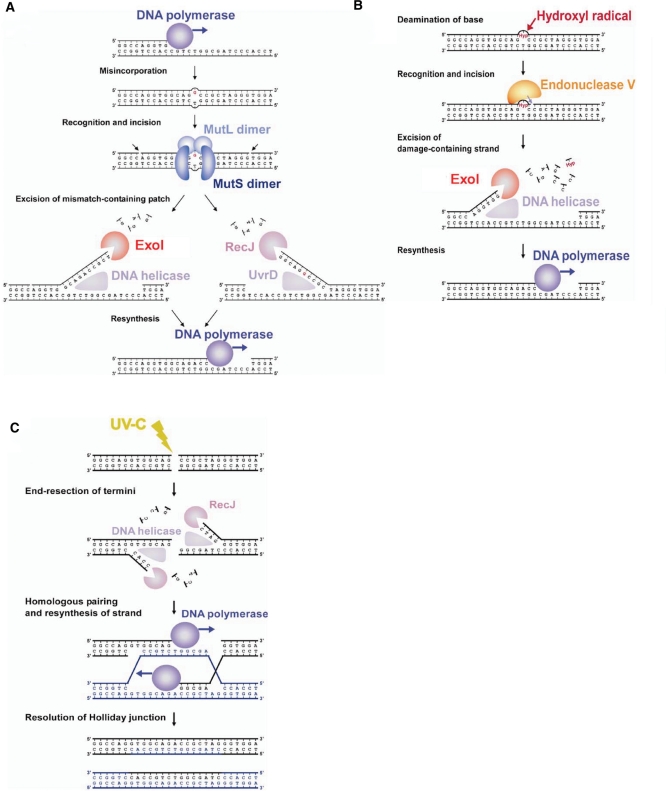
Single-stranded DNA (ssDNA)-specific exonucleases (ssExos) are expected to be involved in a variety of DNA repair pathways corresponding to their cleavage polarities; however, the relationship between the cleavage polarity and the respective DNA repair pathways is only partially understood. To understand the cellular function of ssExos in DNA repair better, genes encoding ssExos were disrupted in Thermus thermophilus HB8 that seems to have only a single set of 5'-3' and 3'-5' ssExos unlike other model organisms. Disruption of the tthb178 gene, which was expected to encode a 3'-5' ssExo, resulted in significant increase in the sensitivity to H(2)O(2) and frequency of the spontaneous mutation rate, but scarcely affected the sensitivity to ultraviolet (UV) irradiation. In contrast, disruption of the recJ gene, which encodes a 5'-3' ssExo, showed little effect on the sensitivity to H(2)O(2), but caused increased sensitivity to UV irradiation. In vitro characterization revealed that TTHB178 possessed 3'-5' ssExo activity that degraded ssDNAs containing deaminated and methylated bases, but not those containing oxidized bases or abasic sites. Consequently, we concluded that TTHB178 is a novel 3'-5' ssExo that functions in various DNA repair systems in cooperation with or independently of RecJ. We named TTHB178 as T. thermophilus exonuclease I.
Figures
Similar articles
-
Overexpression, purification and characterization of RecJ protein from Thermus thermophilus HB8 and its core domain.Nucleic Acids Res. 2001 Nov 15;29(22):4617-24. doi: 10.1093/nar/29.22.4617. Nucleic Acids Res. 2001. PMID: 11713311 Free PMC article.
-
The crystal structure of exonuclease RecJ bound to Mn2+ ion suggests how its characteristic motifs are involved in exonuclease activity.Proc Natl Acad Sci U S A. 2002 Apr 30;99(9):5908-12. doi: 10.1073/pnas.092547099. Epub 2002 Apr 23. Proc Natl Acad Sci U S A. 2002. PMID: 11972066 Free PMC article.
-
Biochemical characterisation of UvrD helicase and RecJ exonuclease from Neisseria gonorrhoeae.Int J Biol Macromol. 2025 May;306(Pt 4):141530. doi: 10.1016/j.ijbiomac.2025.141530. Epub 2025 Mar 1. Int J Biol Macromol. 2025. PMID: 40032130
-
Single-strand DNA-specific exonucleases in Escherichia coli. Roles in repair and mutation avoidance.Genetics. 1998 May;149(1):7-16. doi: 10.1093/genetics/149.1.7. Genetics. 1998. PMID: 9584082 Free PMC article.
-
Structure-specific DNA cleavage by 5' nucleases.Trends Biochem Sci. 1998 Sep;23(9):331-6. doi: 10.1016/s0968-0004(98)01259-6. Trends Biochem Sci. 1998. PMID: 9787638 Review.
Cited by
-
Molecular mechanisms of the whole DNA repair system: a comparison of bacterial and eukaryotic systems.J Nucleic Acids. 2010 Oct 14;2010:179594. doi: 10.4061/2010/179594. J Nucleic Acids. 2010. PMID: 20981145 Free PMC article.
-
Structural insights into the duplex DNA processing of TREX2.Nucleic Acids Res. 2018 Dec 14;46(22):12166-12176. doi: 10.1093/nar/gky970. Nucleic Acids Res. 2018. PMID: 30357414 Free PMC article.
-
DNA mismatch repair in eukaryotes and bacteria.J Nucleic Acids. 2010 Jul 27;2010:260512. doi: 10.4061/2010/260512. J Nucleic Acids. 2010. PMID: 20725617 Free PMC article.
-
Roles of Mn-catalase and a possible heme peroxidase homologue in protection from oxidative stress in Thermus thermophilus.Extremophiles. 2015 Jul;19(4):775-85. doi: 10.1007/s00792-015-0753-2. Epub 2015 May 22. Extremophiles. 2015. PMID: 25997395
-
Structural insights into DNA repair by RNase T--an exonuclease processing 3' end of structured DNA in repair pathways.PLoS Biol. 2014 Mar 4;12(3):e1001803. doi: 10.1371/journal.pbio.1001803. eCollection 2014 Mar. PLoS Biol. 2014. PMID: 24594808 Free PMC article.
References
-
- Crow YJ, Hayward BE, Parmar R, Robins P, Leitch A, Ali M, Black DN, van Bokhoven H, Brunner HG, Hamel BC, et al. Mutations in the gene encoding the 3′-5′ DNA exonuclease TREX1 cause Aicardi-Goutieres syndrome at the AGS1 locus. Nat. Genet. 2006;38:917–920. - PubMed
-
- Lee-Kirsch MA, Gong M, Chowdhury D, Senenko L, Engel K, Lee YA, de Silva U, Bailey SL, Witte T, Vyse TJ, et al. Mutations in the gene encoding the 3′-5′ DNA exonuclease TREX1 are associated with systemic lupus erythematosus. Nat. Genet. 2007;39:1065–1067. - PubMed
-
- Fukuda T, Sumiyoshi T, Takahashi M, Kataoka T, Asahara T, Inui H, Watatani M, Yasutomi M, Kamada N, Miyagawa K. Alterations of the double-strand break repair gene MRE11 in cancer. Cancer Res. 2001;61:23–26. - PubMed
-
- Viswanathan M, Burdett V, Baitinger C, Modrich P, Lovett ST. Redundant exonuclease involvement in Escherichia coli methyl-directed mismatch repair. J. Biol. Chem. 2001;276:31053–31058. - PubMed
-
- Lombardo M-J, Aponyi I, Ray MP, Sandigursky M, Franklin WA, Rosenberg SM. xni-deficient Escherichia coli are proficient for recombination and multiple pathways of repair. DNA Repair. 2003;2:1175–1183. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
