

Botulinum hemagglutinin disrupts the intercellular epithelial barrier by directly binding E-cadherin
- PMID: 20457762
- PMCID: PMC2872904
- DOI: 10.1083/jcb.200910119
Botulinum hemagglutinin disrupts the intercellular epithelial barrier by directly binding E-cadherin
Abstract
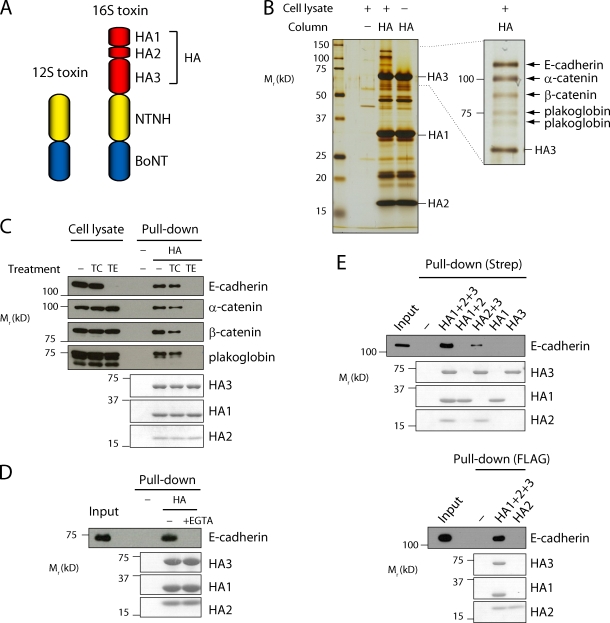
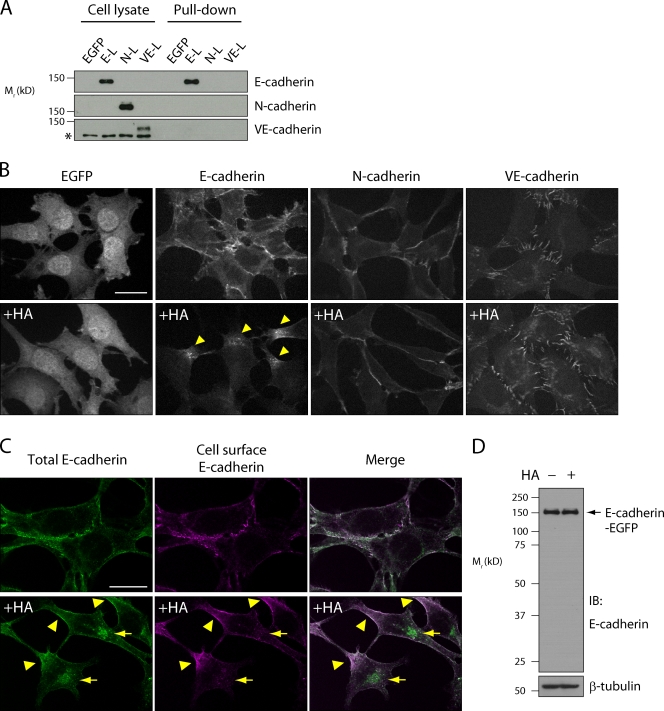
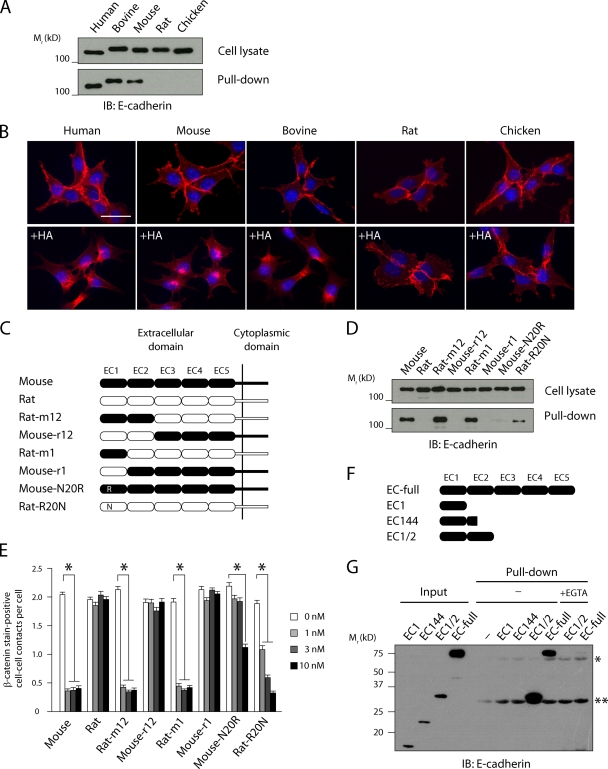
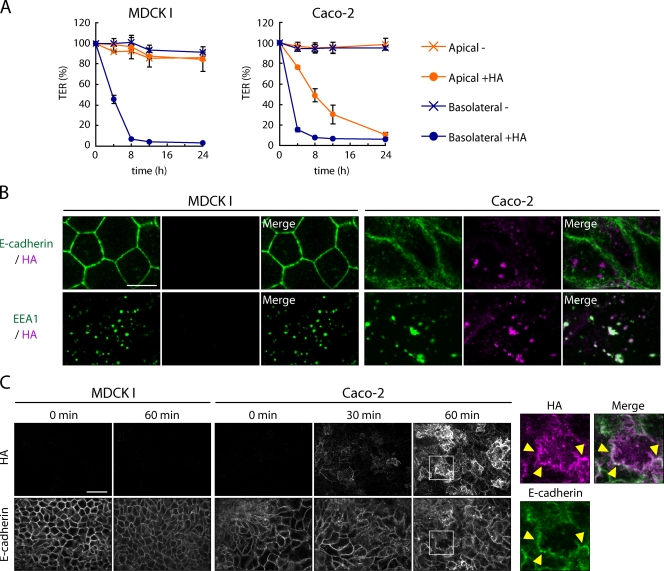
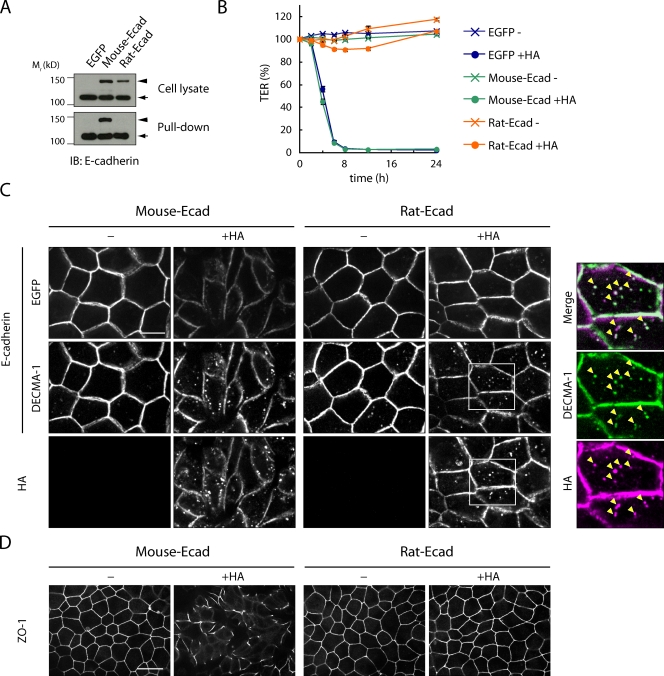
Botulinum neurotoxin is produced by Clostridium botulinum and forms large protein complexes through associations with nontoxic components. We recently found that hemagglutinin (HA), one of the nontoxic components, disrupts the intercellular epithelial barrier; however, the mechanism underlying this phenomenon is not known. In this study, we identified epithelial cadherin (E-cadherin) as a target molecule for HA. HA directly binds E-cadherin and disrupts E-cadherin-mediated cell to cell adhesion. Although HA binds human, bovine, and mouse E-cadherin, it does not bind rat or chicken E-cadherin homologues. HA does not interact with other members of the classical cadherin family such as neural and vascular endothelial cadherin. Expression of rat E-cadherin but not mouse rescues Madin-Darby canine kidney cells from HA-induced tight junction (TJ) disruptions. These data demonstrate that botulinum HA directly binds E-cadherin and disrupts E-cadherin-mediated cell to cell adhesion in a species-specific manner and that the HA-E-cadherin interaction is essential for the disruption of TJ function.
Figures
References
-
- Behrens J., Birchmeier W., Goodman S.L., Imhof B.A. 1985. Dissociation of Madin-Darby canine kidney epithelial cells by the monoclonal antibody anti-arc-1: mechanistic aspects and identification of the antigen as a component related to uvomorulin. J. Cell Biol. 101:1307–1315 10.1083/jcb.101.4.1307 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
