

JIL-1 and Su(var)3-7 interact genetically and counteract each other's effect on position-effect variegation in Drosophila
- PMID: 20457875
- PMCID: PMC2927748
- DOI: 10.1534/genetics.110.117150
JIL-1 and Su(var)3-7 interact genetically and counteract each other's effect on position-effect variegation in Drosophila
Abstract
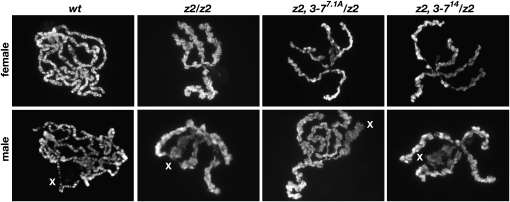
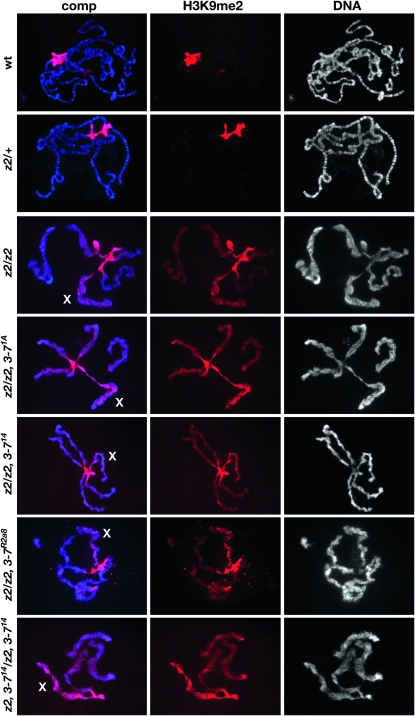
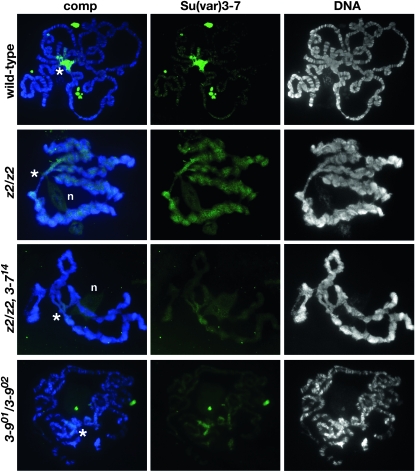
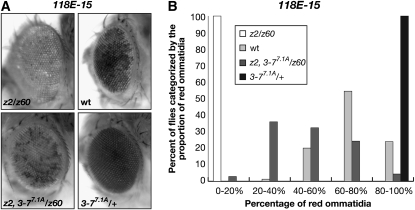
The essential JIL-1 histone H3S10 kinase is a key regulator of chromatin structure that functions to maintain euchromatic domains while counteracting heterochromatization and gene silencing. In the absence of the JIL-1 kinase, two of the major heterochromatin markers H3K9me2 and HP1a spread in tandem to ectopic locations on the chromosome arms. Here we address the role of the third major heterochromatin component, the zinc-finger protein Su(var)3-7. We show that the lethality but not the chromosome morphology defects associated with the null JIL-1 phenotype to a large degree can be rescued by reducing the dose of the Su(var)3-7 gene and that Su(var)3-7 and JIL-1 loss-of-function mutations have an antagonistic and counterbalancing effect on position-effect variegation (PEV). Furthermore, we show that in the absence of JIL-1 kinase activity, Su(var)3-7 gets redistributed and upregulated on the chromosome arms. Reducing the dose of the Su(var)3-7 gene dramatically decreases this redistribution; however, the spreading of H3K9me2 to the chromosome arms was unaffected, strongly indicating that ectopic Su(var)3-9 activity is not a direct cause of lethality. These observations suggest a model where Su(var)3-7 functions as an effector downstream of Su(var)3-9 and H3K9 dimethylation in heterochromatic spreading and gene silencing that is normally counteracted by JIL-1 kinase activity.
Figures
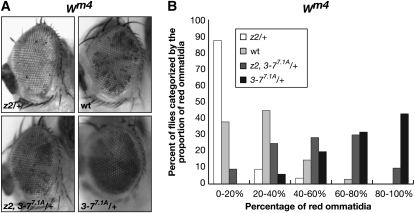
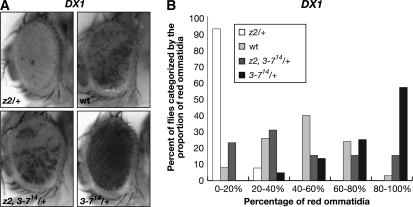
References
-
- Cryderman, D. E., M. H. Cuaycong, S. C. R. Elgin and L. L. Wallrath, 1998. Characterization of sequences associated with position-effect-variegation at pericentric sites in Drosophila heterochromatin. Chromosoma 107 277–285. - PubMed
-
- Danzer, J. R., and L. L. Wallrath, 2004. Mechanisms of HP1-mediated gene silencing in Drosophila. Development 131 3571–3580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
