

Essential role of the Wnt pathway effector Tcf-1 for the establishment of functional CD8 T cell memory
- PMID: 20457902
- PMCID: PMC2906901
- DOI: 10.1073/pnas.0914127107
Essential role of the Wnt pathway effector Tcf-1 for the establishment of functional CD8 T cell memory
Abstract
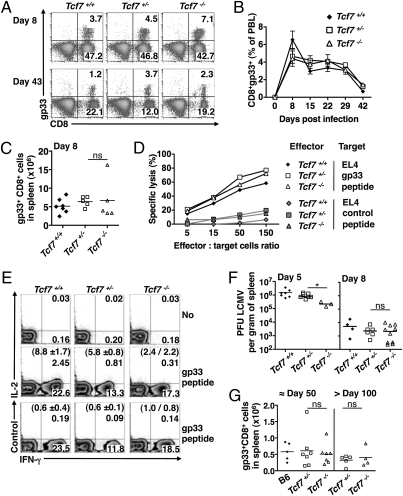
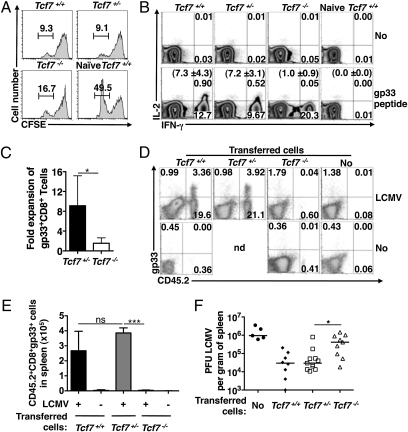
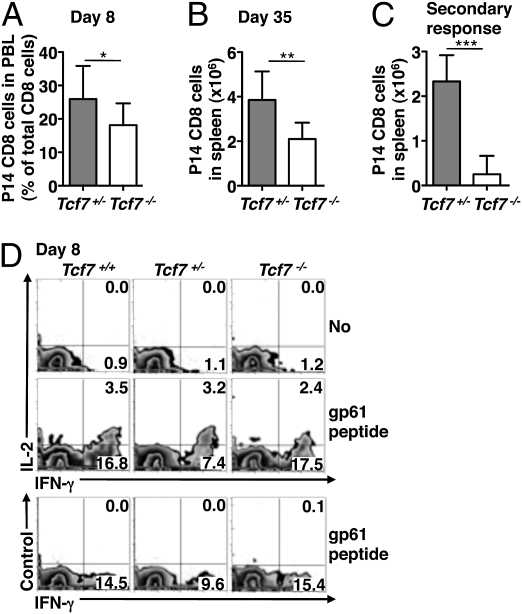
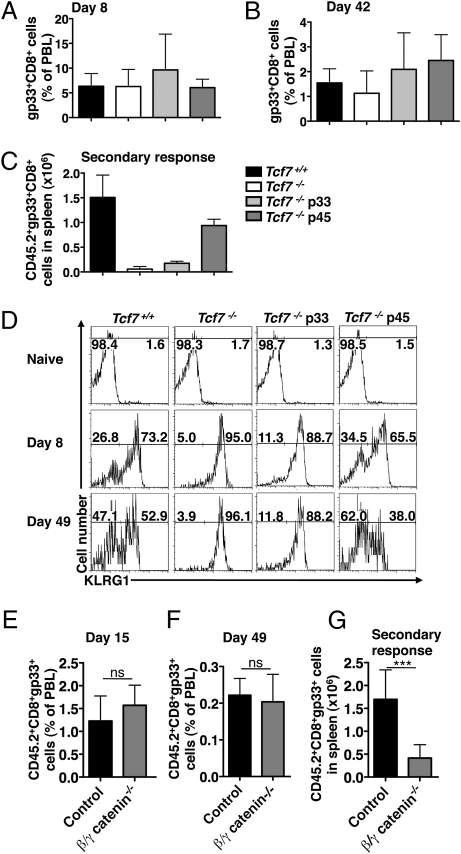
Immune protection from intracellular pathogens depends on the generation of terminally differentiated effector and of multipotent memory precursor CD8 T cells, which rapidly regenerate effector and memory cells during recurrent infection. The identification of factors and pathways involved in CD8 T cell differentiation is of obvious importance to improve vaccination strategies. Here, we show that mice lacking T cell factor 1 (Tcf-1), a nuclear effector of the canonical Wingless/Integration 1 (Wnt) signaling pathway, mount normal effector and effector memory CD8 T cell responses to infection with lymphocytic choriomeningitis virus (LCMV). However, Tcf-1-deficient CD8 T cells are selectively impaired in their ability to expand upon secondary challenge and to protect from recurrent virus infection. Tcf-1-deficient mice essentially lack CD8 memory precursor T cells, which is evident already at the peak of the primary response, suggesting that Tcf-1 programs CD8 memory cell fate. The function of Tcf-1 to establish CD8 T cell memory is dependent on the catenin-binding domain in Tcf-1 and requires the Tcf-1 coactivators and Wnt signaling intermediates beta-catenin and gamma-catenin. These findings demonstrate that the canonical Wnt signaling pathway plays an essential role for CD8 central memory T cell differentiation under physiological conditions in vivo. They raise the possibility that modulation of Wnt signaling may be exploited to improve the generation of CD8 memory T cells during vaccination or for therapies designed to promote sustained cytotoxic CD8 T cell responses against tumors.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Differentiation and persistence of memory CD8(+) T cells depend on T cell factor 1.Immunity. 2010 Aug 27;33(2):229-40. doi: 10.1016/j.immuni.2010.08.002. Immunity. 2010. PMID: 20727791 Free PMC article.
-
Constitutive activation of Wnt signaling favors generation of memory CD8 T cells.J Immunol. 2010 Feb 1;184(3):1191-9. doi: 10.4049/jimmunol.0901199. Epub 2009 Dec 21. J Immunol. 2010. PMID: 20026746 Free PMC article.
-
Differences in the transduction of canonical Wnt signals demarcate effector and memory CD8 T cells with distinct recall proliferation capacity.J Immunol. 2014 Sep 15;193(6):2784-91. doi: 10.4049/jimmunol.1400465. Epub 2014 Aug 15. J Immunol. 2014. PMID: 25127860
-
Interaction of nuclear receptors with the Wnt/beta-catenin/Tcf signaling axis: Wnt you like to know?Endocr Rev. 2005 Dec;26(7):898-915. doi: 10.1210/er.2003-0034. Epub 2005 Aug 26. Endocr Rev. 2005. PMID: 16126938 Review.
-
The canonical Wnt signaling pathway plays an important role in lymphopoiesis and hematopoiesis.Eur J Immunol. 2008 Jul;38(7):1788-94. doi: 10.1002/eji.200738118. Eur J Immunol. 2008. PMID: 18581335 Free PMC article. Review.
Cited by
-
Transcriptional control of effector and memory CD8+ T cell differentiation.Nat Rev Immunol. 2012 Nov;12(11):749-61. doi: 10.1038/nri3307. Epub 2012 Oct 19. Nat Rev Immunol. 2012. PMID: 23080391 Free PMC article. Review.
-
The immunological synapse: the gateway to the HIV reservoir.Immunol Rev. 2013 Jul;254(1):305-25. doi: 10.1111/imr.12080. Immunol Rev. 2013. PMID: 23772628 Free PMC article. Review.
-
Regnase-1 suppresses TCF-1+ precursor exhausted T-cell formation to limit CAR-T-cell responses against ALL.Blood. 2021 Jul 15;138(2):122-135. doi: 10.1182/blood.2020009309. Blood. 2021. PMID: 33690816 Free PMC article.
-
NFκB signaling in T cell memory.Front Immunol. 2023 Feb 24;14:1129191. doi: 10.3389/fimmu.2023.1129191. eCollection 2023. Front Immunol. 2023. PMID: 36911729 Free PMC article. Review.
-
WNT Signaling in Cancer Immunosurveillance.Trends Cell Biol. 2019 Jan;29(1):44-65. doi: 10.1016/j.tcb.2018.08.005. Epub 2018 Sep 13. Trends Cell Biol. 2019. PMID: 30220580 Free PMC article. Review.
References
-
- Stemberger C, et al. Stem cell-like plasticity of naïve and distinct memory CD8+ T cell subsets. Semin Immunol. 2009;21:62–68. - PubMed
-
- Mercado R, et al. Early programming of T cell populations responding to bacterial infection. J Immunol. 2000;165:6833–6839. - PubMed
-
- Stemberger C, et al. A single naive CD8+ T cell precursor can develop into diverse effector and memory subsets. Immunity. 2007;27:985–997. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
