

WD-repeat instability and diversification of the Podospora anserina hnwd non-self recognition gene family
- PMID: 20459612
- PMCID: PMC2873952
- DOI: 10.1186/1471-2148-10-134
WD-repeat instability and diversification of the Podospora anserina hnwd non-self recognition gene family
Abstract
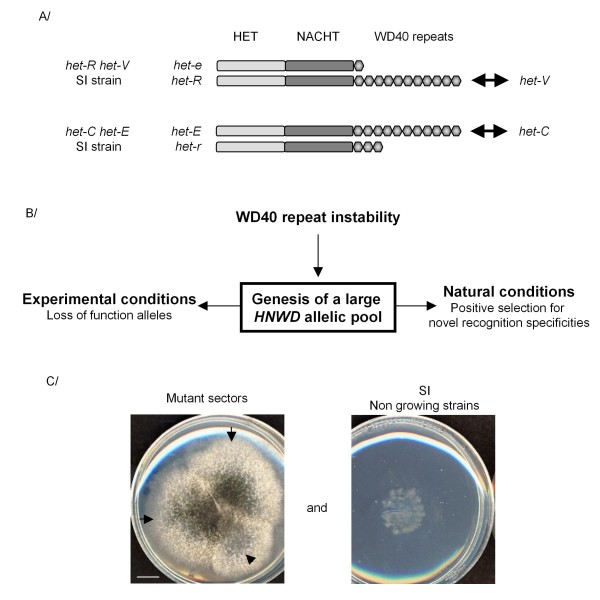
Background: Genes involved in non-self recognition and host defence are typically capable of rapid diversification and exploit specialized genetic mechanism to that end. Fungi display a non-self recognition phenomenon termed heterokaryon incompatibility that operates when cells of unlike genotype fuse and leads to the cell death of the fusion cell. In the fungus Podospora anserina, three genes controlling this allorecognition process het-d, het-e and het-r are paralogs belonging to the same hnwd gene family. HNWD proteins are STAND proteins (signal transduction NTPase with multiple domains) that display a WD-repeat domain controlling recognition specificity. Based on genomic sequence analysis of different P. anserina isolates, it was established that repeat regions of all members of the gene family are extremely polymorphic and undergoing concerted evolution arguing for frequent recombination within and between family members.
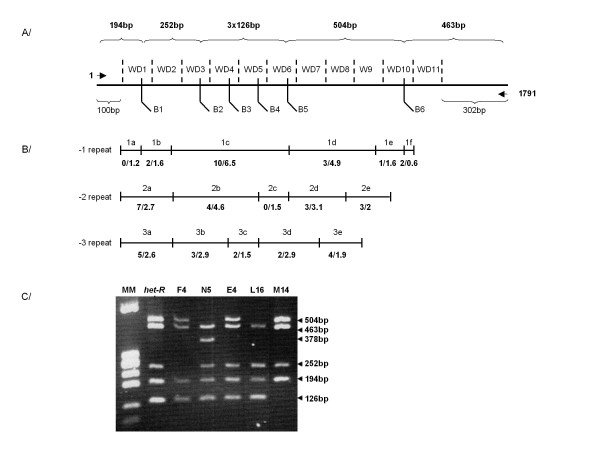
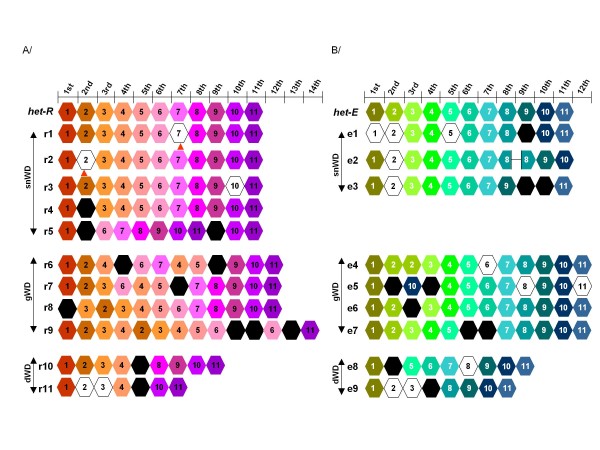
Results: Herein, we directly analyzed the genetic instability and diversification of this allorecognition gene family. We have constituted a collection of 143 spontaneous mutants of the het-R (HNWD2) and het-E (hnwd5) genes with altered recognition specificities. The vast majority of the mutants present rearrangements in the repeat arrays with deletions, duplications and other modifications as well as creation of novel repeat unit variants.
Conclusions: We investigate the extreme genetic instability of these genes and provide a direct illustration of the diversification strategy of this eukaryotic allorecognition gene family.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
