

Sequence features involved in the mechanism of 3' splice junction wobbling
- PMID: 20459675
- PMCID: PMC2875228
- DOI: 10.1186/1471-2199-11-34
Sequence features involved in the mechanism of 3' splice junction wobbling
Abstract
Background: Alternative splicing is an important mechanism mediating the diversified functions of genes in multicellular organisms, and such event occurs in around 40-60% of human genes. Recently, a new splice-junction wobbling mechanism was proposed that subtle modifications exist in mRNA maturation by alternatively choosing at 5'- GTNGT and 3'- NAGNAG, which created single amino acid insertion and deletion isoforms.
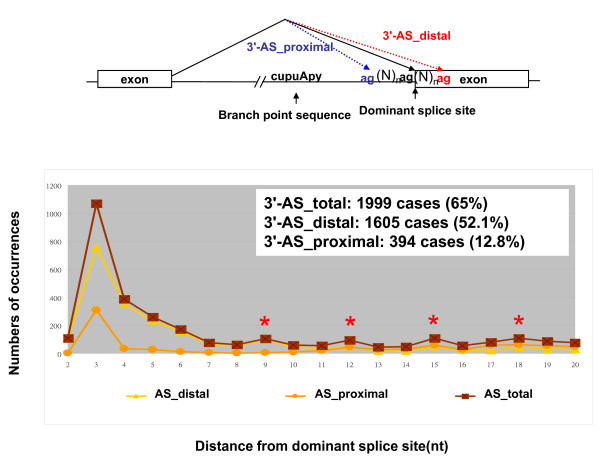
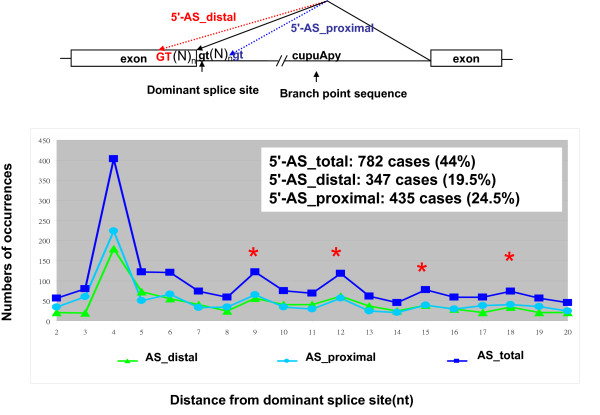
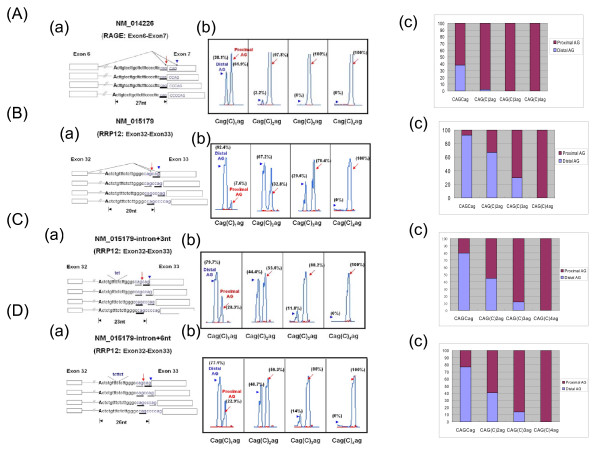
Results: By browsing the Alternative Splicing Database information, we observed that most 3' alternative splice site choices occur within six nucleotides of the dominant splice site and the incidence significantly decreases further away from the dominant acceptor site. Although a lower frequency of alternative splicing occurs within the intronic region (alternative splicing at the proximal AG) than in the exonic region (alternative splicing at the distal AG), alternative AG sites located within the intronic region show stronger potential as the acceptor. These observations revealed that the choice of 3' splice sites during 3' splicing junction wobbling could depend on the distance between the duplicated AG and the branch point site (BPS). Further mutagenesis experiments demonstrated that the distance of AG-to-AG and BPS-to-AG can greatly influence 3' splice site selection. Knocking down a known alternative splicing regulator, hSlu7, failed to affect wobble splicing choices.
Conclusion: Our results implied that nucleotide distance between proximal and distal AG sites has an important regulatory function. In this study, we showed that occurrence of 3' wobble splicing occurs in a distance-dependent manner and that most of this wobble splicing is probably caused by steric hindrance from a factor bound at the neighboring tandem motif sequence.
Figures


References
-
- Sugnet CW, Kent WJ, Ares M, Jr, Haussler D. Transcriptome and genome conservation of alternative splicing events in humans and mice. Pacific Symposium on Biocomputing. 2004. pp. 66–77. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
