

Review
doi: 10.1038/nrm2902.
Epub 2010 May 12.
When pathways collide: collaboration and connivance among signalling proteins in development
Affiliations
- PMID: 20461097
- PMCID: PMC4489880
- DOI: 10.1038/nrm2902
Item in Clipboard
Review
When pathways collide: collaboration and connivance among signalling proteins in development
Nat Rev Mol Cell Biol.
2010 Jun.
Abstract
Signal transduction pathways interact at various levels to define tissue morphology, size and differentiation during development. Understanding the mechanisms by which these pathways collude has been greatly enhanced by recent insights into how shared components are independently regulated and how the activity of one system is contextualized by others. Traditionally, it has been assumed that the components of signalling pathways show pathway fidelity and act with a high degree of autonomy. However, as illustrated by the Wnt and Hippo pathways, there is increasing evidence that components are often shared between multiple pathways and other components talk to each other through multiple mechanisms.
Figures
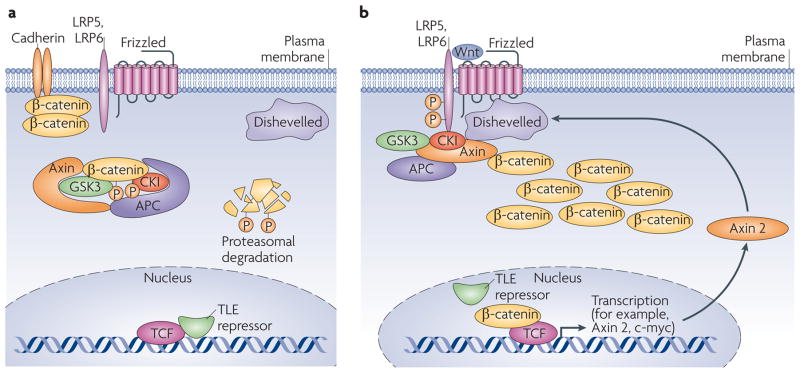
(a) In the absence of a signal, the destruction complex (APC–Axin–GSK-3–CK1) binds and phosphorylates non-cadherin-associated β-catenin, targeting it for destruction by the proteasome. Within the nucleus, TCF/Lef1 DNA-binding proteins are bound by transcriptional repressors (TLE/Groucho). (b) The binding of Wnt ligand to its Frizzled receptor and LRP5/6 co-receptor induces a change in conformation that results in phosphorylation of the co-receptor, which creates a high-affinity binding site for Axin, causing disruption of the destruction complex. This allows β-catenin to accumulate, associate with the TCF/Lef1 proteins, dislodging the TLE repressors and hence promoting transcriptional activation of a program of genes including those encoding c-Myc and Axin2. The latter protein feeds back to inhibit the pathway by promoting assembly of more destruction complexes.
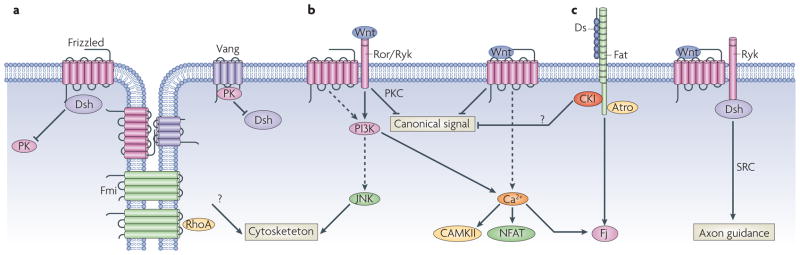
A large number of non-canonical Wnt/PCP signaling pathways have been described. Outlined is a vastly simplified schema highlighting some of the key players. (a) Frizzled-PCP pathway is enriched asymmetrically at cell boundaries. In Drosophila wings Frizzled (Fz), Dishevelled (Dvl) and Diego accumulate at the distal edge of each cells, and Van Gogh (Vang) and Prickle (Pk) accumulate on the proximal side of each cells. These complexes may be bridged by Flamingo-Flamingo (Fmi-Fmi) interactions across cells, as well as interactions between Fmi and Vang and Fmi and Fz and Fz and Vang. In addition there are mutually repressive interactions within the cell between distal and proximal complexes. Genetically RhoA has been placed downstream of the Frizzled-PCP complex. (b) Wnt5a functions primarily as a non-canonical Wnt ligand, acting through the tyrosine kinases Ror or Ryk. Activation of JNK, increases in intracellular calcium, leading to activation of NFAT and inhibition of canonical signaling are some of the alterations seen upon Wnt5a stimulation. (c) Non-canonical Wnts also may function through a subset of Frizzled that bias towards PCP signaling, such as Frizzled7. (d) Dachsous binding to Fat pathway regulates PCP, possibly via recruitment of Casein Kinase I (CKIε), which has been shown in different situations to inhibit or stimulate (not shown) PCP signaling. Fat also recruits the transcriptional co-repressor Atrophin, which suppresses transcription of the PCP effector gene four-jointed (Fj). (e) The atypical tyrosine kinase Ryk is another alternate Wnt receptor, which has been shown to function via Src in axon guidance.
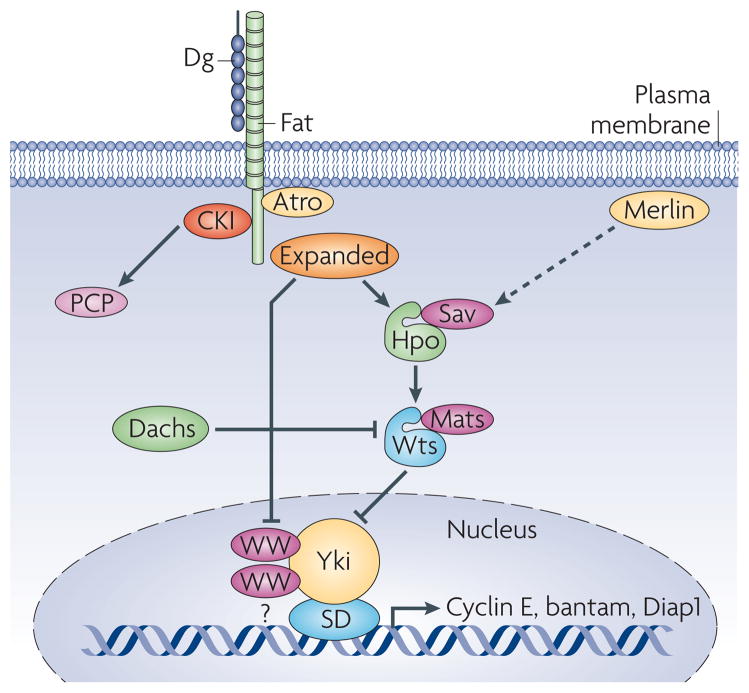
The large cadherin Dachsous (Ds) acts as an inhibitory ligand for the large cadherin Fat. Fat functions to regulate planar cell polarity (PCP) signaling by recruiting the transcriptional co-repressor Atrophin (Atro). Atrophin represses the transcription of the PCP gene four-jointed. It is unclear at present the identity of the transcription factor Atro binds to regulate four-jointed transcription. Fat also regulates growth via control of the Hippo kinase (Hpo) pathway. Current models suggest that Fat regulates Hpo activity either through the FERM domain protein Expanded or the small myosin-like protein Dachs. Dachs has been shown to regulate the stability of Wts Wts phosphorylates Yki, leading to its export from the nucleus. In the absence of Fat or the other Hpo components, Yki promotes the transcription of Cyclin E, the micro RNA bantam and the anti-apoptotic gene Diap1. Sav and Mats are adaptor proteins essential for Hpo and Wts activity. Merlin (Mer) is another FERM domain protein that functions in parallel to Expanded in controlling activity of the Hippo pathway.
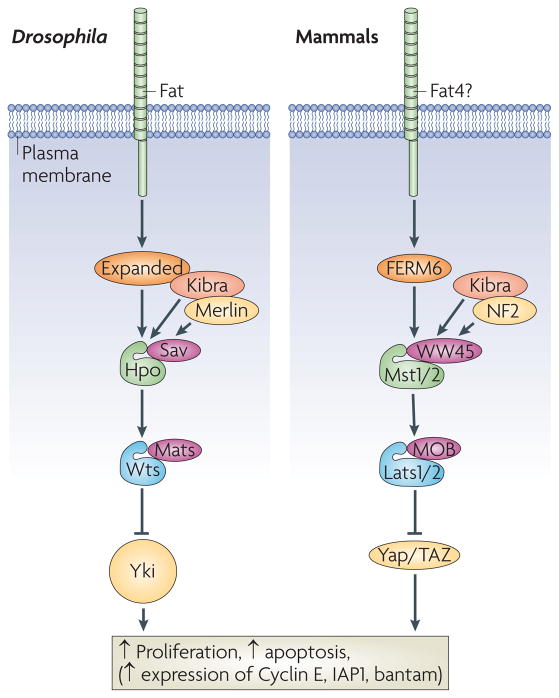
The core Hippo pathway was defined in Drosophila as a cassette composed of the Ste20 family kinase Hippo (Hpo), which forms a complex with the WW domain adaptor protein Salvador (Sav). Hpo phosphorylates and activates the NDR family kinase Warts (Wts), which is complexed with the adaptor protein MATS (‘Mob as Tumour Suppressor’). Wts phosphorylates the transcriptional co-activator Yorkie (Yki). Phosphorylated Yki is bound by 14-3-3 proteins and excluded from the nucleus. When in the nucleus, Yki promotes the transcription of growth-promoting and apoptosis-inhibiting genes, such as Cyclin E and Diap1. Upstream of Hpo lies the cadherin Fat, and the FERM domain proteins Expanded (Ex) and Merlin. Kibra is a WW domain protein that promotes Ex-Mer interactions, and enhances Hpo pathway activity. The analogous pathway in mammals is outlined on the right. YAP and TAZ are homologous to Yorkie, Mst1/2 are homologous to Hpo, Lats1/2 are homologous to Wts, Sav is homologous to WW45, NF2 is homologous to Merlin and FRMD6 is homologous to Ex. The vertebrate homolog of Fat is Fat4, but this protein has not yet been shown to function in the vertebrate Hippo pathway.
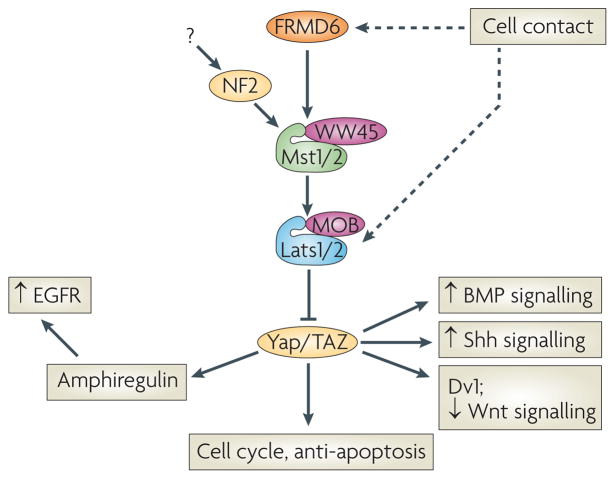
Recent studies have uncovered unsuspected cross-talk between the Hippo pathway and other growth regulators. Cell contact is still detected in MEFs lacking both Mts1 and Mts2, indicating an Mst-independent input (possibly a kinase) that can phosphorylate and activate Lats1 and Lats2. Lats1/2 then inhibits YAP/TAZ to control growth. YAP can function as a transcriptional co-activator of SMADs, enhancing BMP signaling. YAP also can stimulate Gli transcription, enhancing Shh signaling. AKT can inhibit Mst1 kinase, providing another avenue for growth promotion and anti-apoptotic activity of PI3K/AKT signaling pathways. TAZ can bind Dishevelled (Dvl), inhibiting β-catenin signaling in the cytoplasm. When TAZ translocates to the nucleus, it frees Dvl to activate β-catenin, enhancing Wnt signaling. Nuclear YAP also regulates the expression of Amphiregulin, an EGFR ligand. Therefore loss of Mst or Lats in one cell can drive proliferation in that cell, while simultaneously activating the proliferation of nearby cells via the EGFR pathway.
References
-
- Cohen P. The role of protein phosphorylation in neural and hormonal control of cellular activity. Nature. 1982;296:613–20. - PubMed
-
- van Amerongen R, Nusse R. Towards an integrated view of Wnt signaling in development. Development. 2009;136:3205–14. This reference and the one following (3) provide excellent and up to date reviews of Wnt signaling. - PubMed
-
- Zeng Q, Hong W. The emerging role of the hippo pathway in cell contact inhibition, organ size control, and cancer development in mammals. Cancer Cell. 2008;13:188–92. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
