

doi: 10.1186/1759-8753-1-15.
Non-long terminal repeat (non-LTR) retrotransposons: mechanisms, recent developments, and unanswered questions
Affiliations
- PMID: 20462415
- PMCID: PMC2881922
- DOI: 10.1186/1759-8753-1-15
Item in Clipboard
Non-long terminal repeat (non-LTR) retrotransposons: mechanisms, recent developments, and unanswered questions
Mob DNA.
.
Abstract
Non-long terminal repeat (non-LTR) retrotransposons are present in most eukaryotic genomes. In some species, such as humans, these elements are the most abundant genome sequence and continue to replicate to this day, creating a source of endogenous mutations and potential genotoxic stress. This review will provide a general outline of the replicative cycle of non-LTR retrotransposons. Recent findings regarding the host regulation of non-LTR retrotransposons will be summarized. Finally, future directions of interest will be discussed.
Figures
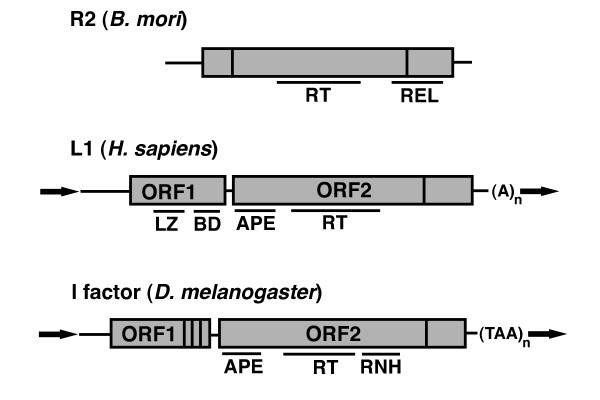
Structure of non-long terminal repeat (non-LTR) retrotransposons. The general structures of three model non-LTR elements are shown. Top: the Bombyx mori R2 element consists of a single open reading frame (ORF) containing reverse transcriptase (RT) activity and a restriction enzyme-like (REL) domain. Vertical lines indicate cysteine-histidine rich regions believed to encode nucleic acid binding domains. Horizontal lines are untranslated regions. Middle: the human L1 element contains two ORFs. ORF1 contains a leucine zipper (LZ) domain involved in protein-protein interactions and a C-terminal nucleic acid binding domain (BD). ORF2 has a N-terminal apurinic/apyrimidinic endonuclease (APE), a central RT domain, and a C-terminal cysteine-histidine rich domain. The element ends with a polyA tail. Typical insertions are flanked by target site duplications (bold arrows). Bottom: the Drosophila I factor element. ORF1 contains three C-terminal cysteine-histidine rich motifs resembling those of retroviral gag. ORF2 contains an APE endonuclease, central RT/RNaseH (RNH) domain, and C-terminal cysteine-histidine domain. In all structures contiguous gray boxes represent a single ORF.
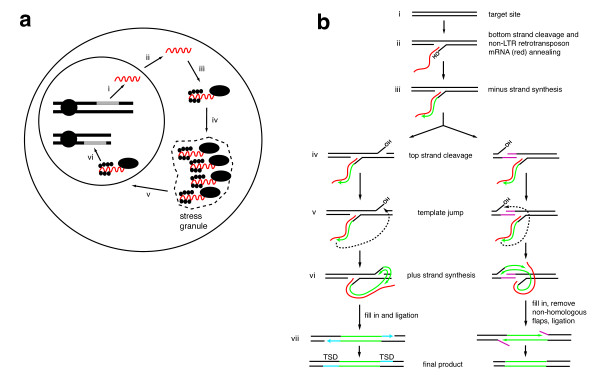
Replication of non-long terminal repeat (non-LTR) retrotransposons. (a) Replicative cycle of non-LTR retrotransposition. (i) Transcription of a full-length, active element. (ii) mRNA export from the nucleus. (iii) Translation of retrotransposon proteins and (iv) passage through cytoplasmic granule. (v) Ribonucleoprotein particle (RNP) import into the nucleus. (vi) Integration via target-primed reverse transcription (TPRT). (b) A model for TPRT. (i) Original unmodified target site. This sequence at this site will vary depending on the specificity of the retrotransposon endonuclease. (ii) Cleavage of one strand of the target site (bottom strand in the figure) by endonuclease. (iii) Minus strand synthesis using retrotransposon mRNA as a template. (iv) During or after minus strand synthesis, top strand cleavage occurs by the retrotransposon endonuclease or a cellular endonuclease. A downstream cleavage (left pathway) leads to a target site duplication (TSD). An upstream cleavage (right pathway) leads to a target site deletion. (v) Template jump from mRNA to top strand of target site. (vi) Plus strand synthesis using the cleaved top strand as primer. (vii) Left pathway: complete of synthesis and fill in of gaps (blue) leads to a target site duplication. Right pathway: completion of synthesis and degradation of non-homologous flaps by unknown nucleases leads to a target site deletion (purple sequences). There are alternative products that are not shown in this model for simplicity.
References
-
- Dawkins R. The Selfish Gene. UK Oxford University Press; 1976.
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, Funke R, Gage D, Harris K, Heaford A, Howland J, Kann L, Lehoczky J, LeVine R, McEwan P, McKernan K, Meldrim J, Mesirov JP, Miranda C, Morris W, Naylor J, Raymond C, Rosetti M, Santos R, Sheridan A, Sougnez C. Initial sequencing and analysis of the human genome. Nature. 2001;409:860–921. doi: 10.1038/35057062. - DOI - PubMed
-
- Mouse Genome Sequencing Consortium; Waterston RH, Lindblad-Toh K, Birney E, Rogers J, Abril JF, Agarwal P, Agarwala R, Ainscough R, Alexandersson M, An P, Antonarakis SE, Attwood J, Baertsch R, Bailey J, Barlow K, Beck S, Berry E, Birren B, Bloom T, Bork P, Botcherby M, Bray N, Brent MR, Brown DG, Brown SD, Bult C, Burton J, Butler J, Campbell RD. Initial sequencing and comparative analysis of the mouse genome. Nature. 2002;420:520–562. doi: 10.1038/nature01262. - DOI - PubMed
-
- Schnable PS, Ware D, Fulton RS, Stein JC, Wei F, Pasternak S, Liang C, Zhang J, Fulton L, Graves TA, Minx P, Reily AD, Courtney L, Kruchowski SS, Tomlinson C, Strong C, Delehaunty K, Fronick C, Courtney B, Rock SM, Belter E, Du F, Kim K, Abbott RM, Cotton M, Levy A, Marchetto P, Ochoa K, Jackson SM, Gillam B. The B73 maize genome: complexity, diversity, and dynamics. Science. 2009;326:1112–1115. doi: 10.1126/science.1178534. - DOI - PubMed
-
- Goff SA, Ricke D, Lan TH, Presting G, Wang R, Dunn M, Glazebrook J, Sessions A, Oeller P, Varma H, Hadley D, Hutchison D, Martin C, Katagiri F, Lange BM, Moughamer T, Xia Y, Budworth P, Zhong J, Miguel T, Paszkowski U, Zhang S, Colbert M, Sun WL, Chen L, Cooper B, Park S, Wood TC, Mao L, Quail P. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica) Science. 2002;296:92–100. doi: 10.1126/science.1068275. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
