

Ubr1 and Ubr2 function in a quality control pathway for degradation of unfolded cytosolic proteins
- PMID: 20462952
- PMCID: PMC2893976
- DOI: 10.1091/mbc.e10-02-0098
Ubr1 and Ubr2 function in a quality control pathway for degradation of unfolded cytosolic proteins
Abstract
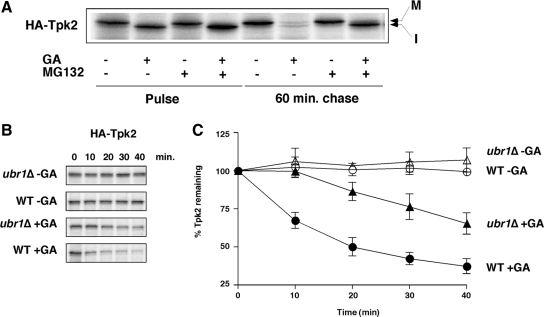
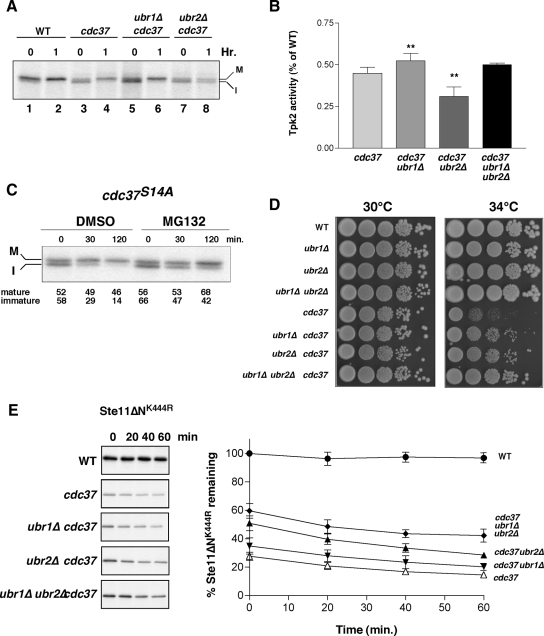
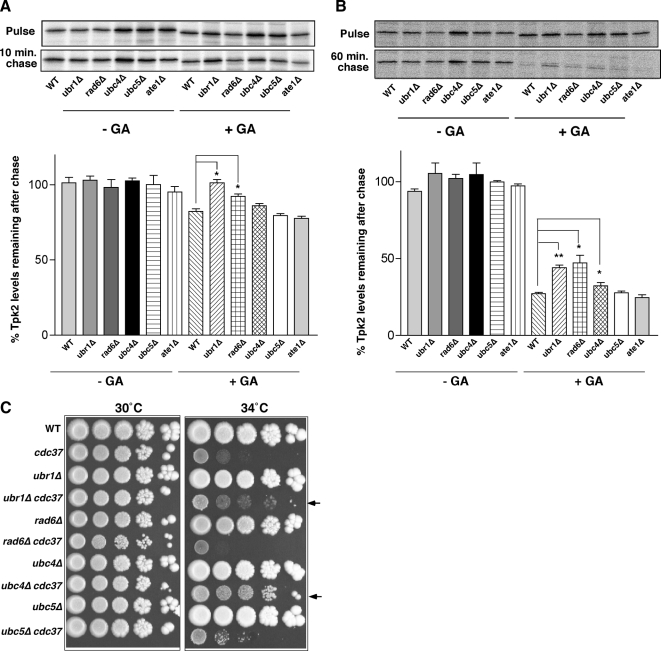
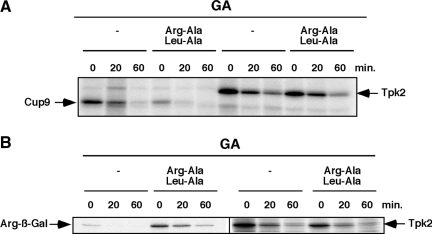
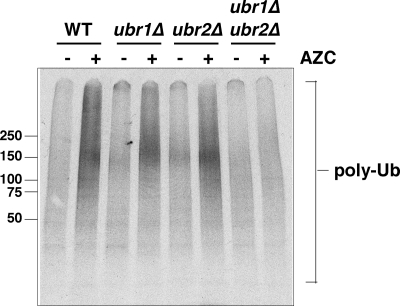
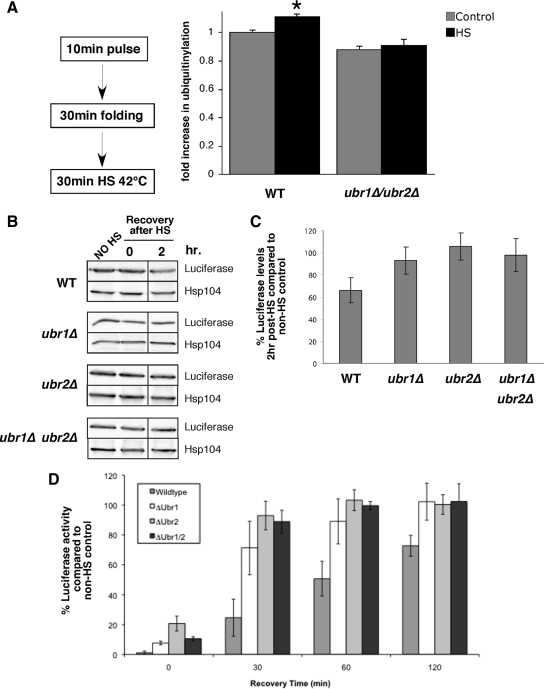
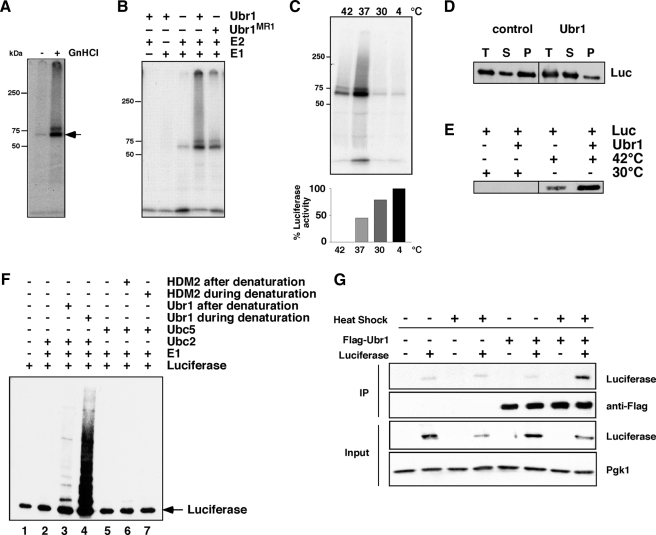
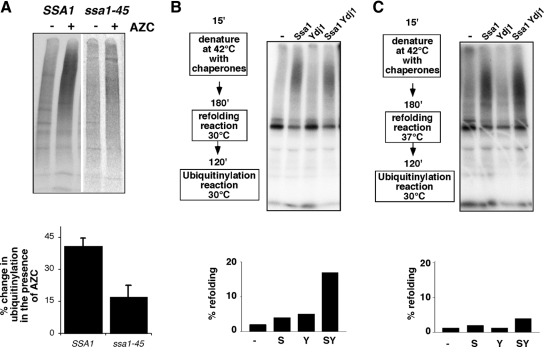
Quality control systems facilitate polypeptide folding and degradation to maintain protein homeostasis. Molecular chaperones promote folding, whereas the ubiquitin/proteasome system mediates degradation. We show here that Saccharomyces cerevisiae Ubr1 and Ubr2 ubiquitin ligases promote degradation of unfolded or misfolded cytosolic polypeptides. Ubr1 also catalyzes ubiquitinylation of denatured but not native luciferase in a purified system. This activity is based on the direct interaction of denatured luciferase with Ubr1, although Hsp70 stimulates polyubiquitinylation of the denatured substrate. We also report that loss of Ubr1 and Ubr2 function suppressed the growth arrest phenotype resulting from chaperone mutation. This correlates with increased protein kinase maturation and indicates partitioning of foldable conformers toward the proteasome. Our findings, based on the efficiency of this quality control system, suggest that the cell trades growth potential to avert the potential toxicity associated with accumulation of unfolded or misfolded proteins. Ubr1 and Ubr2 therefore represent E3 components of a novel quality control pathway for proteins synthesized on cytosolic ribosomes.
Figures
Similar articles
-
The Type II Hsp40 Sis1 cooperates with Hsp70 and the E3 ligase Ubr1 to promote degradation of terminally misfolded cytosolic protein.PLoS One. 2013;8(1):e52099. doi: 10.1371/journal.pone.0052099. Epub 2013 Jan 16. PLoS One. 2013. PMID: 23341891 Free PMC article.
-
Direct involvement of Hsp70 ATP hydrolysis in Ubr1-dependent quality control.Mol Biol Cell. 2020 Nov 15;31(24):2669-2686. doi: 10.1091/mbc.E20-08-0541. Epub 2020 Sep 23. Mol Biol Cell. 2020. PMID: 32966159 Free PMC article.
-
A network of ubiquitin ligases is important for the dynamics of misfolded protein aggregates in yeast.J Biol Chem. 2012 Jul 6;287(28):23911-22. doi: 10.1074/jbc.M112.341164. Epub 2012 May 16. J Biol Chem. 2012. PMID: 22593585 Free PMC article.
-
Hul5 ubiquitin ligase: good riddance to bad proteins.Prion. 2012 Jul 1;6(3):240-4. doi: 10.4161/pri.19929. Epub 2012 Jul 1. Prion. 2012. PMID: 22561164 Free PMC article. Review.
-
False start: cotranslational protein ubiquitination and cytosolic protein quality control.J Proteomics. 2014 Apr 4;100:92-101. doi: 10.1016/j.jprot.2013.08.005. Epub 2013 Aug 15. J Proteomics. 2014. PMID: 23954725 Review.
Cited by
-
It's all about talking: two-way communication between proteasomal and lysosomal degradation pathways via ubiquitin.Am J Physiol Cell Physiol. 2016 Aug 1;311(2):C166-78. doi: 10.1152/ajpcell.00074.2016. Epub 2016 May 25. Am J Physiol Cell Physiol. 2016. PMID: 27225656 Free PMC article.
-
Hsp70 clears misfolded kinases that partitioned into distinct quality-control compartments.Mol Biol Cell. 2015 May 1;26(9):1583-600. doi: 10.1091/mbc.E14-08-1262. Epub 2015 Mar 4. Mol Biol Cell. 2015. PMID: 25739454 Free PMC article.
-
Biology of the heat shock response and protein chaperones: budding yeast (Saccharomyces cerevisiae) as a model system.Microbiol Mol Biol Rev. 2012 Jun;76(2):115-58. doi: 10.1128/MMBR.05018-11. Microbiol Mol Biol Rev. 2012. PMID: 22688810 Free PMC article. Review.
-
Studying Spatial Protein Quality Control, Proteopathies, and Aging Using Different Model Misfolding Proteins in S. cerevisiae.Front Mol Neurosci. 2018 Jul 23;11:249. doi: 10.3389/fnmol.2018.00249. eCollection 2018. Front Mol Neurosci. 2018. PMID: 30083092 Free PMC article. Review.
-
Ubiquitin ligase ITCH recruitment suppresses the aggregation and cellular toxicity of cytoplasmic misfolded proteins.Sci Rep. 2014 May 28;4:5077. doi: 10.1038/srep05077. Sci Rep. 2014. PMID: 24865853 Free PMC article.
References
-
- Balch W. E., Morimoto R. I., Dillin A., Kelly J. W. Adapting proteostasis for disease intervention. Science. 2008;319:916–919. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
