

TBC-2 regulates RAB-5/RAB-7-mediated endosomal trafficking in Caenorhabditis elegans
- PMID: 20462958
- PMCID: PMC2893991
- DOI: 10.1091/mbc.e09-11-0947
TBC-2 regulates RAB-5/RAB-7-mediated endosomal trafficking in Caenorhabditis elegans
Abstract
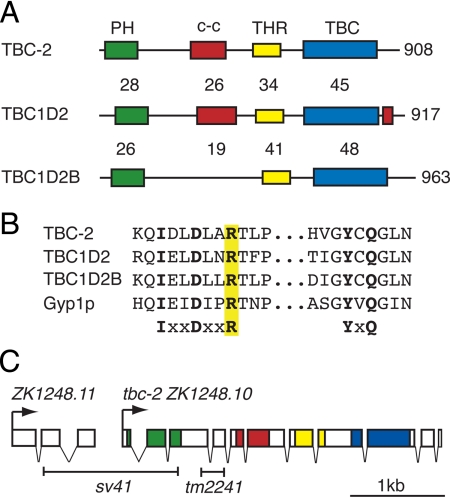
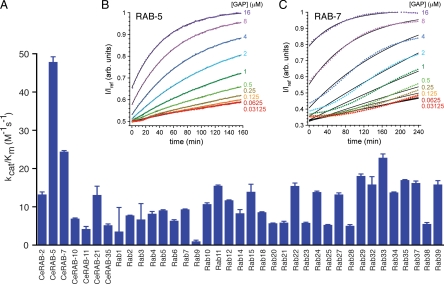
During endosome maturation the early endosomal Rab5 GTPase is replaced with the late endosomal Rab7 GTPase. It has been proposed that active Rab5 can recruit and activate Rab7, which in turn could inactivate and remove Rab5. However, many of the Rab5 and Rab7 regulators that mediate endosome maturation are not known. Here, we identify Caenorhabditis elegans TBC-2, a conserved putative Rab GTPase-activating protein (GAP), as a regulator of endosome to lysosome trafficking in several tissues. We show that tbc-2 mutant animals accumulate enormous RAB-7-positive late endosomes in the intestine containing refractile material. RAB-5, RAB-7, and components of the homotypic fusion and vacuole protein sorting (HOPS) complex, a RAB-7 effector/putative guanine nucleotide exchange factor (GEF), are required for the tbc-2(-) intestinal phenotype. Expression of activated RAB-5 Q78L in the intestine phenocopies the tbc-2(-) large late endosome phenotype in a RAB-7 and HOPS complex-dependent manner. TBC-2 requires the catalytic arginine-finger for function in vivo and displays the strongest GAP activity on RAB-5 in vitro. However, TBC-2 colocalizes primarily with RAB-7 on late endosomes and requires RAB-7 for membrane localization. Our data suggest that TBC-2 functions on late endosomes to inactivate RAB-5 during endosome maturation.
Figures







Similar articles
-
The VPS34 PI3K negatively regulates RAB-5 during endosome maturation.J Cell Sci. 2017 Jun 15;130(12):2007-2017. doi: 10.1242/jcs.194746. Epub 2017 Apr 28. J Cell Sci. 2017. PMID: 28455411 Free PMC article.
-
Identification of the switch in early-to-late endosome transition.Cell. 2010 Apr 30;141(3):497-508. doi: 10.1016/j.cell.2010.03.011. Cell. 2010. PMID: 20434987
-
Basolateral Endocytic Recycling Requires RAB-10 and AMPH-1 Mediated Recruitment of RAB-5 GAP TBC-2 to Endosomes.PLoS Genet. 2015 Sep 22;11(9):e1005514. doi: 10.1371/journal.pgen.1005514. eCollection 2015. PLoS Genet. 2015. PMID: 26393361 Free PMC article.
-
Who's in control? Principles of Rab GTPase activation in endolysosomal membrane trafficking and beyond.J Cell Biol. 2021 Sep 6;220(9):e202105120. doi: 10.1083/jcb.202105120. Epub 2021 Aug 12. J Cell Biol. 2021. PMID: 34383013 Free PMC article. Review.
-
Vps34 and the Armus/TBC-2 Rab GAPs: Putting the brakes on the endosomal Rab5 and Rab7 GTPases.Cell Logist. 2017 Dec 19;7(4):e1403530. doi: 10.1080/21592799.2017.1403530. eCollection 2017. Cell Logist. 2017. PMID: 29296513 Free PMC article. Review.
Cited by
-
Evi5 promotes collective cell migration through its Rab-GAP activity.J Cell Biol. 2012 Jul 9;198(1):57-67. doi: 10.1083/jcb.201112114. J Cell Biol. 2012. PMID: 22778279 Free PMC article.
-
DRG2 is required for surface localization of PD-L1 and the efficacy of anti-PD-1 therapy.Cell Death Discov. 2024 May 27;10(1):260. doi: 10.1038/s41420-024-02027-x. Cell Death Discov. 2024. PMID: 38802348 Free PMC article.
-
TBC1D9B functions as a GTPase-activating protein for Rab11a in polarized MDCK cells.Mol Biol Cell. 2014 Nov 15;25(23):3779-97. doi: 10.1091/mbc.E13-10-0604. Epub 2014 Sep 17. Mol Biol Cell. 2014. PMID: 25232007 Free PMC article.
-
The conserved SNARE SEC-22 localizes to late endosomes and negatively regulates RNA interference in Caenorhabditis elegans.RNA. 2017 Mar;23(3):297-307. doi: 10.1261/rna.058438.116. Epub 2016 Dec 14. RNA. 2017. PMID: 27974622 Free PMC article.
-
The VPS34 PI3K negatively regulates RAB-5 during endosome maturation.J Cell Sci. 2017 Jun 15;130(12):2007-2017. doi: 10.1242/jcs.194746. Epub 2017 Apr 28. J Cell Sci. 2017. PMID: 28455411 Free PMC article.
References
-
- Albert S., Gallwitz D. Two new members of a family of Ypt/Rab GTPase activating proteins. Promiscuity of substrate recognition. J. Biol. Chem. 1999;274:33186–33189. - PubMed
-
- Bernards A. GAPs galore! A survey of putative Ras superfamily GTPase activating proteins in man and Drosophila. Biochim. Biophys. Acta. 2003;1603:47–82. - PubMed
-
- Brune M., Hunter J. L., Corrie J. E., Webb M. R. Direct, real-time measurement of rapid inorganic phosphate release using a novel fluorescent probe and its application to actomyosin subfragment 1 ATPase. Biochemistry. 1994;33:8262–8271. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
