

The eukaryotic initiation factor (eIF) 4G HEAT domain promotes translation re-initiation in yeast both dependent on and independent of eIF4A mRNA helicase
- PMID: 20463023
- PMCID: PMC2903371
- DOI: 10.1074/jbc.M110.132027
The eukaryotic initiation factor (eIF) 4G HEAT domain promotes translation re-initiation in yeast both dependent on and independent of eIF4A mRNA helicase
Abstract
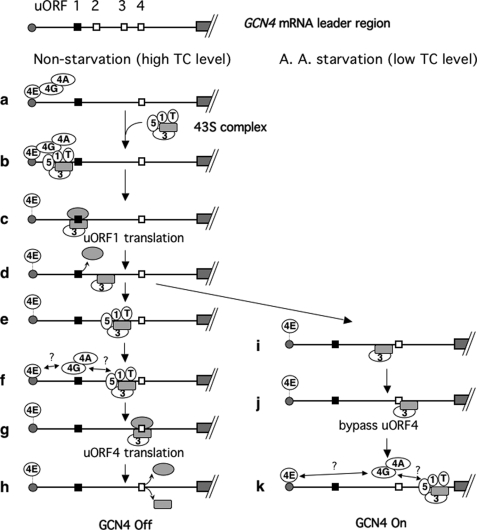
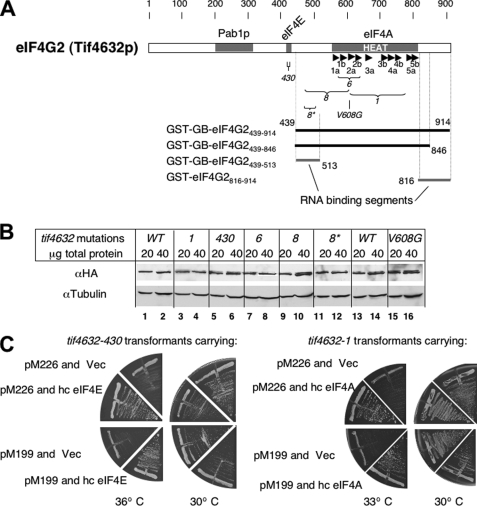
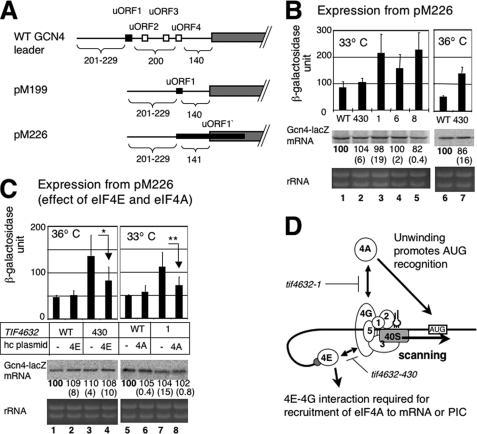
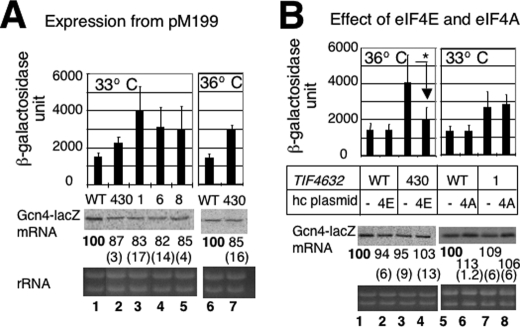
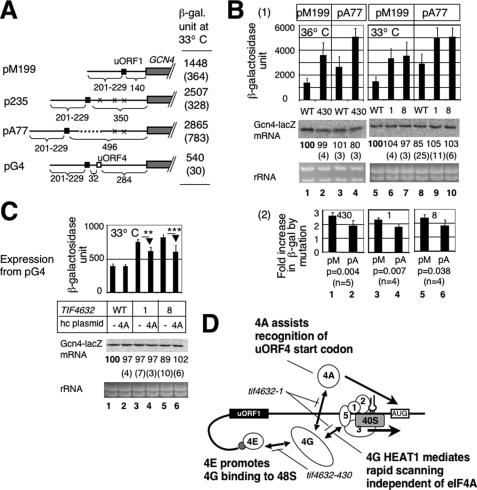
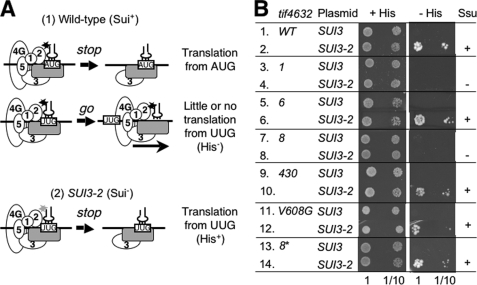
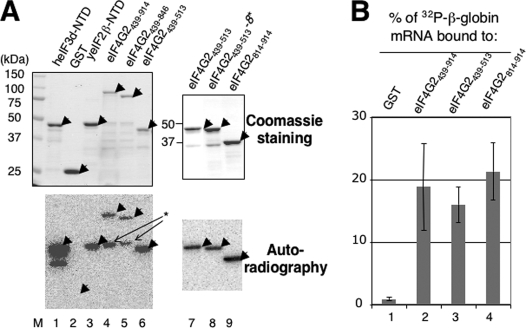
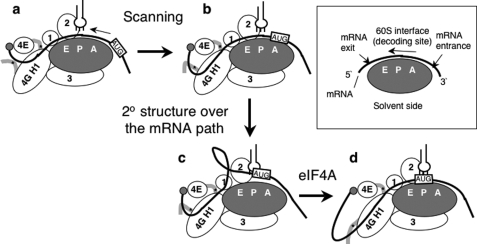
Translation re-initiation provides the molecular basis for translational control of mammalian ATF4 and yeast GCN4 mediated by short upstream open reading (uORFs) in response to eIF2 phosphorylation. eIF4G is the major adaptor subunit of eIF4F that binds the cap-binding subunit eIF4E and the mRNA helicase eIF4A and is also required for re-initiation in mammals. Here we show that the yeast eIF4G2 mutations altering eIF4E- and eIF4A-binding sites increase re-initiation at GCN4 and impair recognition of the start codons of uORF1 or uORF4 located after uORF1. The increase in re-initiation at GCN4 was partially suppressed by increasing the distance between uORF1 and GCN4, suggesting that the mutations decrease the migration rate of the scanning ribosome in the GCN4 leader. Interestingly, eIF4E overexpression suppressed both the phenotypes caused by the mutation altering eIF4E-binding site. Thus, eIF4F is required for accurate AUG selection and re-initiation also in yeast, and the eIF4G interaction with the mRNA-cap appears to promote eIF4F re-acquisition by the re-initiating 40 S subunit. However, eIF4A overexpression suppressed the impaired AUG recognition but not the increase in re-initiation caused by the mutations altering eIF4A-binding site. These results not only provide evidence that mRNA unwinding by eIF4A stimulates start codon recognition, but also suggest that the eIF4A-binding site on eIF4G made of the HEAT domain stimulates the ribosomal scanning independent of eIF4A. Based on the RNA-binding activities identified within the unstructured segments flanking the eIF4G2 HEAT domain, we discuss the role of the HEAT domain in scanning beyond loading eIF4A onto the pre-initiation complex.
Figures
Similar articles
-
Yeast eukaryotic initiation factor 4B (eIF4B) enhances complex assembly between eIF4A and eIF4G in vivo.J Biol Chem. 2013 Jan 25;288(4):2340-54. doi: 10.1074/jbc.M112.398537. Epub 2012 Nov 26. J Biol Chem. 2013. PMID: 23184954 Free PMC article.
-
Translation initiation factors GleIF4E2 and GleIF4A can interact directly with the components of the pre-initiation complex to facilitate translation initiation in Giardia lamblia.Mol Biochem Parasitol. 2020 Mar;236:111258. doi: 10.1016/j.molbiopara.2020.111258. Epub 2020 Jan 20. Mol Biochem Parasitol. 2020. PMID: 31968220
-
The domains of yeast eIF4G, eIF4E and the cap fine-tune eIF4A activities through an intricate network of stimulatory and inhibitory effects.Nucleic Acids Res. 2022 Jun 24;50(11):6497-6510. doi: 10.1093/nar/gkac437. Nucleic Acids Res. 2022. PMID: 35689631 Free PMC article.
-
Gene-specific translational control of the yeast GCN4 gene by phosphorylation of eukaryotic initiation factor 2.Mol Microbiol. 1993 Oct;10(2):215-23. doi: 10.1111/j.1365-2958.1993.tb01947.x. Mol Microbiol. 1993. PMID: 7934812 Review.
-
Manipulation of the host translation initiation complex eIF4F by DNA viruses.Biochem Soc Trans. 2010 Dec;38(6):1511-6. doi: 10.1042/BST0381511. Biochem Soc Trans. 2010. PMID: 21118117 Review.
Cited by
-
DDX3 regulates endoplasmic reticulum stress-induced ATF4 expression.Sci Rep. 2017 Oct 23;7(1):13832. doi: 10.1038/s41598-017-14262-7. Sci Rep. 2017. PMID: 29062139 Free PMC article.
-
A mechanistic overview of translation initiation in eukaryotes.Nat Struct Mol Biol. 2012 Jun 5;19(6):568-76. doi: 10.1038/nsmb.2303. Nat Struct Mol Biol. 2012. PMID: 22664984 Review.
-
Interaction between 25S rRNA A loop and eukaryotic translation initiation factor 5B promotes subunit joining and ensures stringent AUG selection.Mol Cell Biol. 2013 Sep;33(18):3540-8. doi: 10.1128/MCB.00771-13. Epub 2013 Jul 8. Mol Cell Biol. 2013. PMID: 23836883 Free PMC article.
-
Characterization of the role of eIF4G in stimulating cap- and IRES-dependent translation in aplysia neurons.PLoS One. 2013 Sep 3;8(9):e74085. doi: 10.1371/journal.pone.0074085. eCollection 2013. PLoS One. 2013. PMID: 24019950 Free PMC article.
-
Analysing GCN4 translational control in yeast by stochastic chemical kinetics modelling and simulation.BMC Syst Biol. 2011 Aug 18;5:131. doi: 10.1186/1752-0509-5-131. BMC Syst Biol. 2011. PMID: 21851603 Free PMC article.
References
-
- Hinnebusch A. G., Dever T. E., Asano K. (2007) in Translational Control in Biology and Medicine (Mathews M. B., Sonenberg N., Hershey J. W. eds) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY: 225–268
-
- Pestova T. V., Lorsch J. R., Hellen C. U. (2007) in Translational Control in Biology and Medicine (Mathews M. B., Sonenberg N., Hershey J. W. eds) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY: 87–128
-
- Asano K., Sachs M. S. (2007) Genes Dev. 21, 1280–1287 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
