

Guidance of postural motoneurons requires MAPK/ERK signaling downstream of fibroblast growth factor receptor 1
- PMID: 20463222
- PMCID: PMC6632572
- DOI: 10.1523/JNEUROSCI.4932-09.2010
Guidance of postural motoneurons requires MAPK/ERK signaling downstream of fibroblast growth factor receptor 1
Abstract
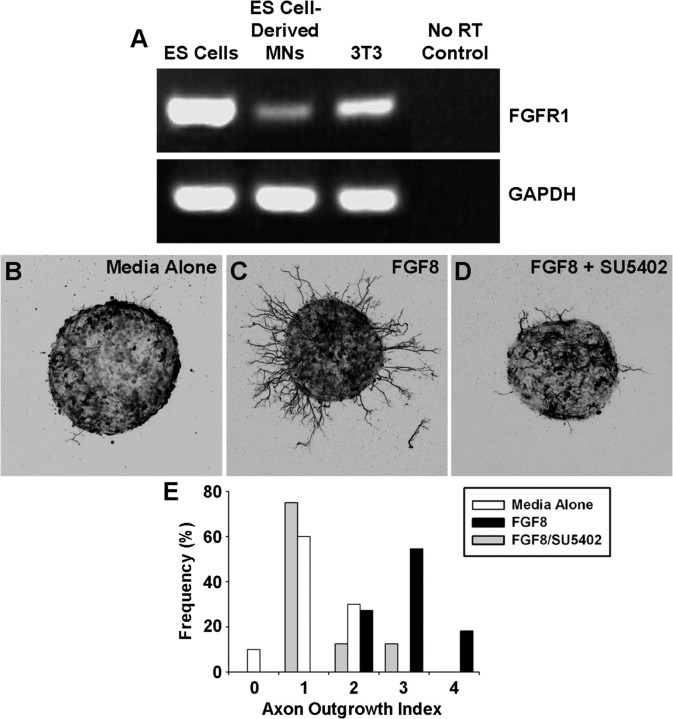
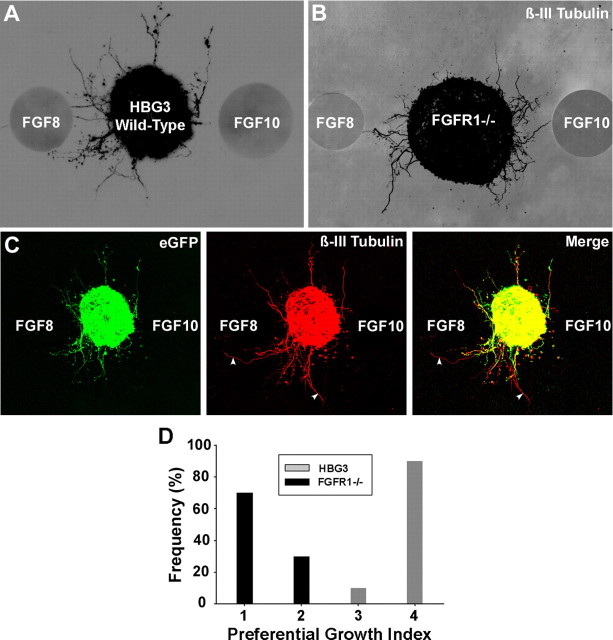
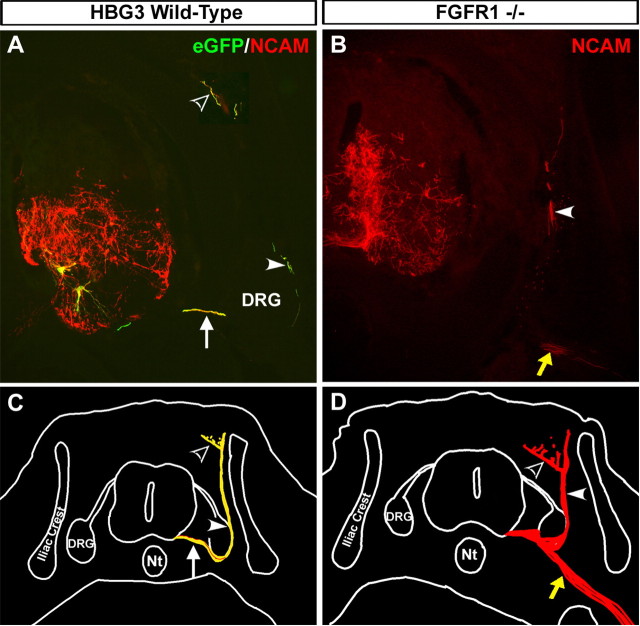
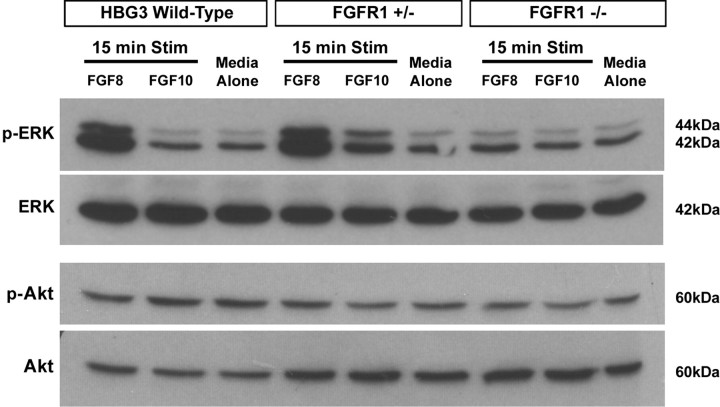
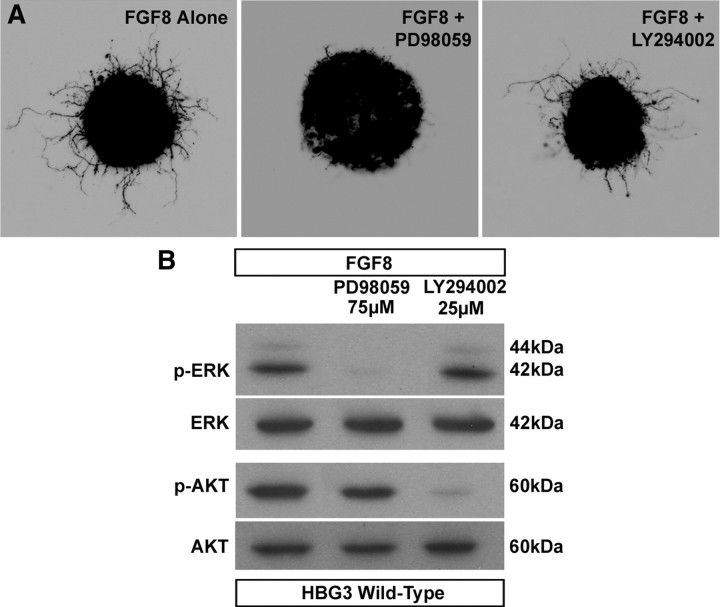
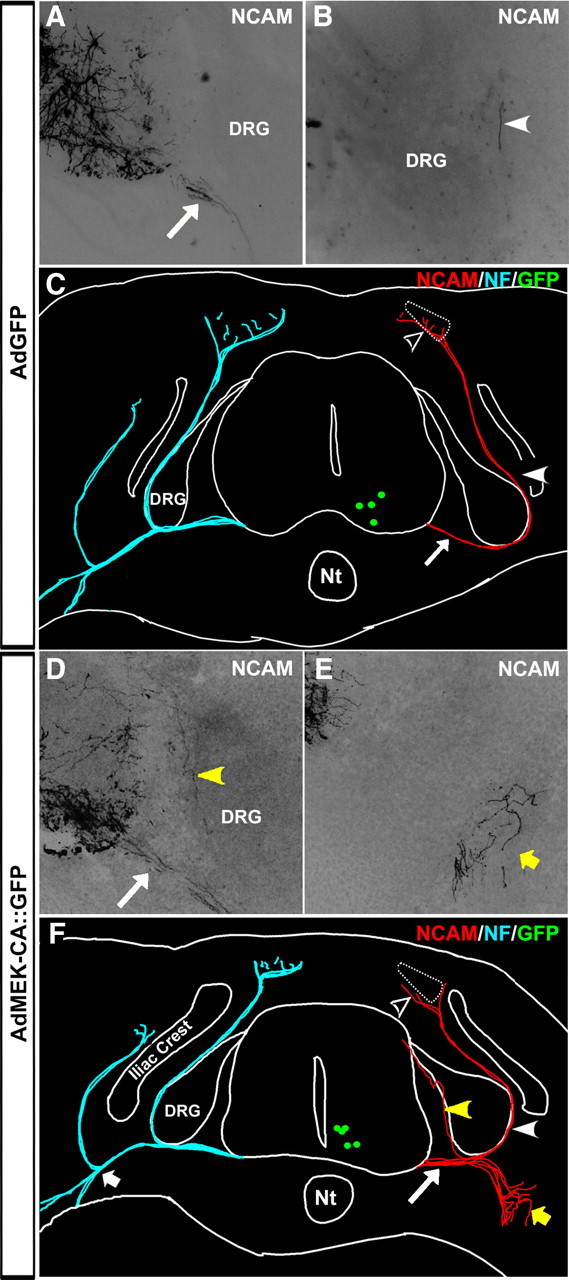
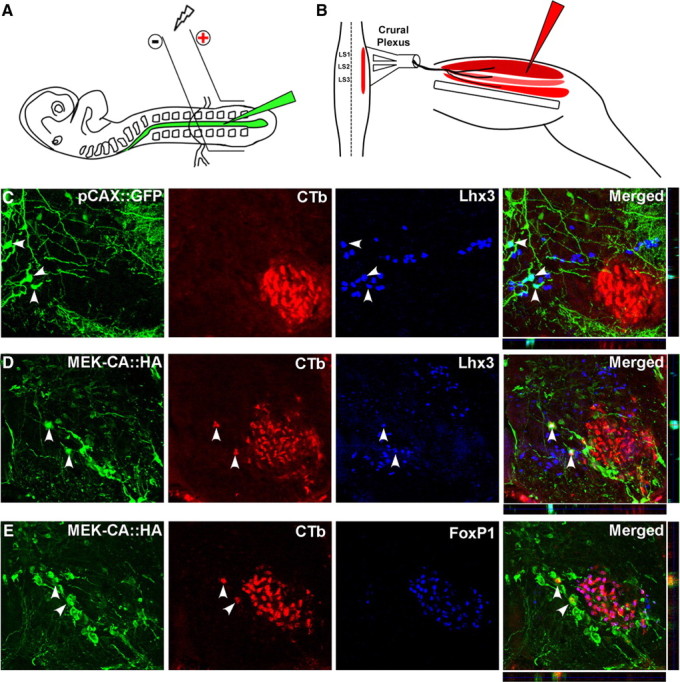
Identification of intracellular signaling pathways necessary for appropriate axon guidance is challenging because many CNS populations used to study these events contain multiple cell types. Here, we resolve this issue by using mouse embryonic stem (ES) cells that were directed to differentiate into a population of motoneurons that exclusively innervate epaxial muscles [medial median motor column (MMCm) motoneurons]. These ES cell-derived MMCm motoneurons, like their endogenous counterparts, express fibroblast growth factor receptor 1 (FGFR1) and selectively extend axons toward the epaxial trophin FGF8. Unlike wild-type MMCm motoneurons, FGFR1(-/-) MMCm motoneurons show guidance defects when transplanted into the neural tube of chick embryos. Furthermore, activation of FGFR1 selectively signals through mitogen-activated protein kinase/extracellular signal-regulated kinase (MAPK/ERK) for appropriate guidance in vitro, whereas overexpression of constitutively active MAPK/ERK in transplanted, or endogenous chick, MMCm cells causes guidance defects in vivo. These results indicate that MAPK/ERK activation downstream of FGFR1 is necessary for MMCm motor axon guidance and that ES cell-derived neurons provide an important tool for dissecting intracellular pathways required for axon guidance.
Figures
Similar articles
-
Motoneurons derived from embryonic stem cells express transcription factors and develop phenotypes characteristic of medial motor column neurons.J Neurosci. 2006 Mar 22;26(12):3256-68. doi: 10.1523/JNEUROSCI.5537-05.2006. J Neurosci. 2006. PMID: 16554476 Free PMC article.
-
FGF as a target-derived chemoattractant for developing motor axons genetically programmed by the LIM code.Neuron. 2006 Jun 15;50(6):841-53. doi: 10.1016/j.neuron.2006.04.030. Neuron. 2006. PMID: 16772167
-
Endocytosis and Transport of Growth Factor Receptors in Peripheral Axon Regeneration: Novel Lessons from Neurons Expressing Lysine-Deficient FGF Receptor Type 1 in vitro.Anat Rec (Hoboken). 2019 Aug;302(8):1268-1275. doi: 10.1002/ar.24120. Epub 2019 Apr 22. Anat Rec (Hoboken). 2019. PMID: 30950230 Free PMC article.
-
Spontaneous rhythmic activity in early chick spinal cord influences distinct motor axon pathfinding decisions.Brain Res Rev. 2008 Jan;57(1):77-85. doi: 10.1016/j.brainresrev.2007.06.021. Epub 2007 Aug 1. Brain Res Rev. 2008. PMID: 17920131 Free PMC article. Review.
-
Development of neuromuscular connections: guidance of motoneuron axons to muscles in the embryonic chick hindlimb.Ciba Found Symp. 1988;138:97-115. doi: 10.1002/9780470513675.ch7. Ciba Found Symp. 1988. PMID: 3058436 Review.
Cited by
-
[Relationship between FGFR1 Gene Regulation of Circulating Tumor Cells and Clinical Features of Non-small Cell Lung Cancer].Zhongguo Fei Ai Za Zhi. 2018 May 20;21(5):365-374. doi: 10.3779/j.issn.1009-3419.2018.05.03. Zhongguo Fei Ai Za Zhi. 2018. PMID: 29764586 Free PMC article. Chinese.
-
Motor neurons and the generation of spinal motor neuron diversity.Front Cell Neurosci. 2014 Oct 9;8:293. doi: 10.3389/fncel.2014.00293. eCollection 2014. Front Cell Neurosci. 2014. PMID: 25346659 Free PMC article. Review.
-
Non-Mammalian Models for Understanding Neurological Defects in RASopathies.Biomedicines. 2024 Apr 10;12(4):841. doi: 10.3390/biomedicines12040841. Biomedicines. 2024. PMID: 38672195 Free PMC article. Review.
-
Zebrafish rgs4 is essential for motility and axonogenesis mediated by Akt signaling.Cell Mol Life Sci. 2013 Mar;70(5):935-50. doi: 10.1007/s00018-012-1178-z. Epub 2012 Oct 11. Cell Mol Life Sci. 2013. PMID: 23052218 Free PMC article.
-
Inhibition of fibroblast growth factor receptor signaling impairs metastasis of hepatocellular carcinoma.Tumour Biol. 2014 Nov;35(11):11005-11. doi: 10.1007/s13277-014-2384-0. Epub 2014 Aug 5. Tumour Biol. 2014. Retraction in: Tumour Biol. 2017 Apr 20. doi: 10.1007/s13277-017-5487-6. PMID: 25091573 Retracted.
References
-
- Atwal JK, Massie B, Miller FD, Kaplan DR. The TrkB-Shc site signals neuronal survival and local axon growth via MEK and P13-kinase. Neuron. 2000;27:265–277. - PubMed
-
- Barres BA, Silverstein BE, Corey DP, Chun LL. Immunological, morphological, and electrophysiological variation among retinal ganglion cells purified by panning. Neuron. 1988;1:791–803. - PubMed
-
- Campbell DS, Holt CE. Apoptotic pathway and MAPKs differentially regulate chemotropic responses of retinal growth cones. Neuron. 2003;37:939–952. - PubMed
-
- Colamarino SA, Tessier-Lavigne M. The axonal chemoattractant netrin-1 is also a chemorepellent for trochlear motor axons. Cell. 1995;81:621–629. - PubMed
-
- Colvin JS, Bohne BA, Harding GW, McEwen DG, Ornitz DM. Skeletal overgrowth and deafness in mice lacking fibroblast growth factor receptor 3. Nat Genet. 1996;12:390–397. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous