

doi: 10.1523/JNEUROSCI.5420-09.2010.
Evidence for the involvement of Lfc and Tctex-1 in axon formation
Affiliations
- PMID: 20463241
- PMCID: PMC3845415
- DOI: 10.1523/JNEUROSCI.5420-09.2010
Item in Clipboard
Evidence for the involvement of Lfc and Tctex-1 in axon formation
J Neurosci.
.
Abstract
RhoA and Rac play key and opposite roles during neuronal polarization. We now show that Lfc, a guanosine nucleotide exchange factor (GEF), localizes to the Golgi apparatus and growth cones of developing neurons and negatively regulates neurite sprouting and axon formation through a Rho signaling pathway. Tctex-1, a dynein light chain implicated in axon outgrowth by modulating actin dynamics and Rac activity, colocalizes and physically interacts with Lfc, thus inhibiting its GEF activity, decreasing Rho-GTP levels, and functionally antagonizing Lfc during neurite formation.
Figures
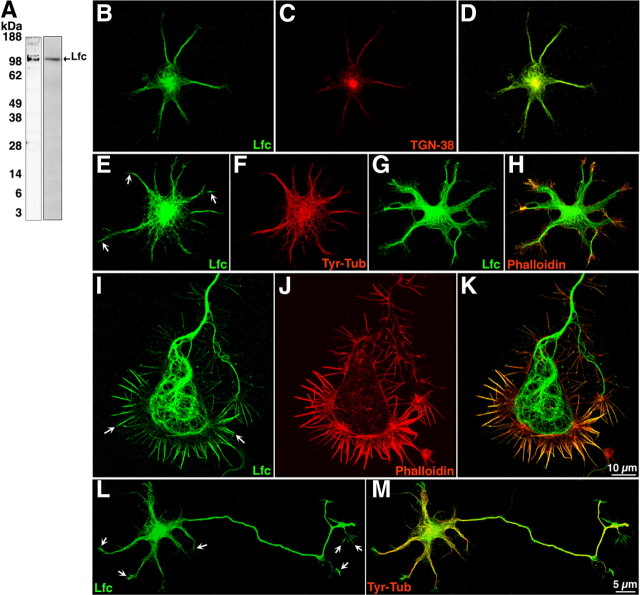
Lfc subcellular distribution in young cultured hippocampal pyramidal neurons. A, Western blots showing specificity of the polyclonal antibodies used for detecting Lfc. Left lane, Chicken polyclonal antibody; right lane, rabbit polyclonal antibody. Both antibodies were probed by immunoblotting a brain lysate obtained from a postnatal rat brain. B–D, Confocal images showing colocalization of Lfc (green) and TGN-38 (red) in the Golgi apparatus. E–H, Confocal images showing stage 2 neurons stained for Lfc (green) and tyr-tubulin (F, red), or rhodamine–phalloidin (H, red). Note that Lfc immunolabeling is concentrated in the cell center, where it colocalizes with the Golgi marker TGN38, associates with microtubules, and extends into the microtubule-free region at neuritic tips (E, arrows) where it localizes with F-actin. I–K, A high-power view of a large axonal growth cone from a stage 2–3 neuron showing the distribution of Lfc (green) and rhodamine–phalloidin (red). Note that Lfc associates with microtubules located in the growth cone C domain and with actin-rich filopodial extensions located in the growth cone P domain (arrows). The transition zone between the C and P domains displays little Lfc immunolabeling. L, M, Confocal images showing a stage 3 neuron immunostained for Lfc (green) and tyr-tubulin (red). Note that Lfc immunolabeling is present in all types of neurites (minor process or the axon) and that it extends into neuritic tips (arrows). Also note that Lfc immunolabeling is higher in the axonal shaft than in minor neurites. Images of Lfc immunostaining were obtained using the rabbit polyclonal antibody; similar staining patterns were observed with the chicken polyclonal antibody.
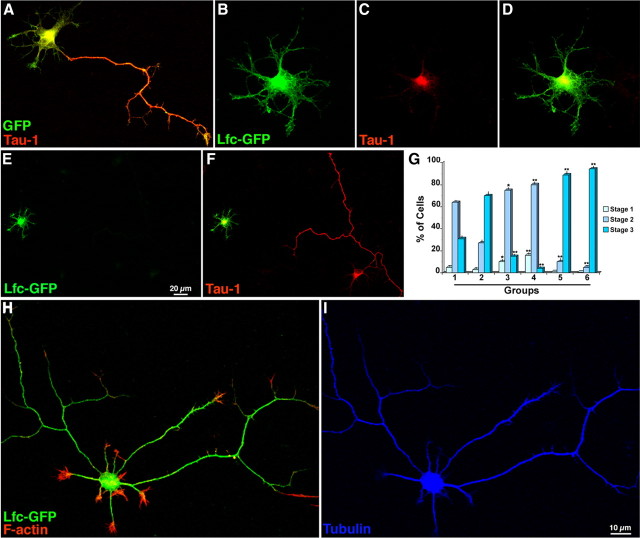
Lfc overexpression inhibits axon formation. A, Confocal image showing a cultured hippocampal neuron transfected with GFP (green) 2 h after plating, fixed 24 h later, and stained with a mAb against Tau (Tau-1; red). B–D, A similar experiment, but from a culture transfected with Lfc-GFP. Note that the transfected cell is arrested at stage 2 of neuritic development; also note that Tau immunolabeling (C) is restricted to the cell body. E, F, Another example of a neuron transfected with Lfc-GFP that has failed to develop an axon; in contrast, a nontransfected neuron present in the same field has extended a single long and branched Tau+ (F, red) axon. G, Graph showing the percentage of cells at stages 1–3 of neuritic development after transfection with either GFP or Lfc-GFP; cultures were transfected 2 h after plating and fixed 24 or 30 h later. The different groups represent the following: 1, GFP (24 h); 2, GFP (30 h); 3, Lfc-GFP (24 h); 4, Lfc-GFP (30 h); 5, Lfc-GFP plus Y27632 (24 h); 6, Lfc-GFP plus Y27632 (30 h). Note that Lfc overexpression significantly (*p < 0.05; **p < 0.001) increases the number of cells at stages 1 and 2 while decreasing those at stage 3 of neuritic development. Also note that Y27632 reverts the inhibitory effect of Lfc and significantly (**p < 0.001) increases the percentage of cells at stage 3. H, I, Confocal images showing the morphology of an Lfc-GFP-transfected neuron (H, green) treated with Y27632; the inhibitor (37 nmol/ml) was added to the culture medium 2 h after transfection, fixed 24 h later, and labeled for tyr-tubulin (blue) and rhodamine-phalloidin (red). Note that the Lfc-GFP-transfected neuron has extended several long axon-like neurites; a similar response was observed in control GFP-transfected neurons (data not shown).
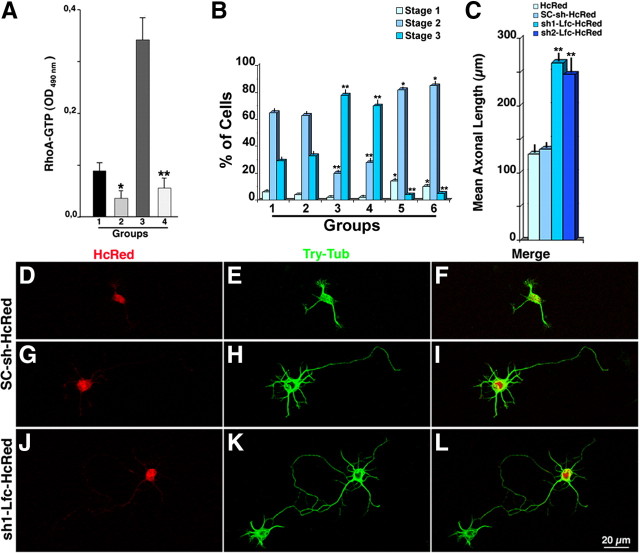
A, Graph showing levels of endogenous RhoA-GTP in SC-sh-HcRed-transfected (1) and sh1-Lfc-HcRed-transfected (2) hippocampal pyramidal neurons. It also shows Rho-GTP levels after transfection with Lfc-GFP (3) and Lfc-GFP plus sh1-Lfc-HcRed (4). Note that Lfc suppression significantly (*p < 0.05; **p < 0.001) decreases endogenous and Lfc-induced RhoA GTP activity. B, Graph showing the percentage of cells at stages 1–3 of neuritic development after transfection with HcRed (1) SC-sh-HcRed (2), sh1-Lfc-HcRed (3), sh2-Lfc-HcRed (4), constitutively active RhoA (5), or sh1-Lfc-HcRed plus constitutively active RhoA (6). Cultures were transfected 2 h after plating and fixed 22 h later. All plasmids were used at a concentration of 2 μg/ml. Values are significantly different from those of the corresponding control groups: *p < 0.05; **p < 0.001. C, Graph showing that Lfc suppression increases the length of axonal processes. Neurons were transfected 2 h after plating and fixed at the end of the first day in vitro. They were stained with the mAb Tau-1, and the length of axonal processes were measured using MetaMorph software. Note that suppression of Lfc significantly (**p < 0.001) increases axonal length. D–L, Confocal images showing the morphology of cultured hippocampal pyramidal neurons after transfection with SC-sh-HcRed and sh1-Lfc-HcRed. Neurons were transfected 2 h after plating, fixed at the end of the first day in vitro, and labeled with a mAb against tyrosinated α-tubulin (green).
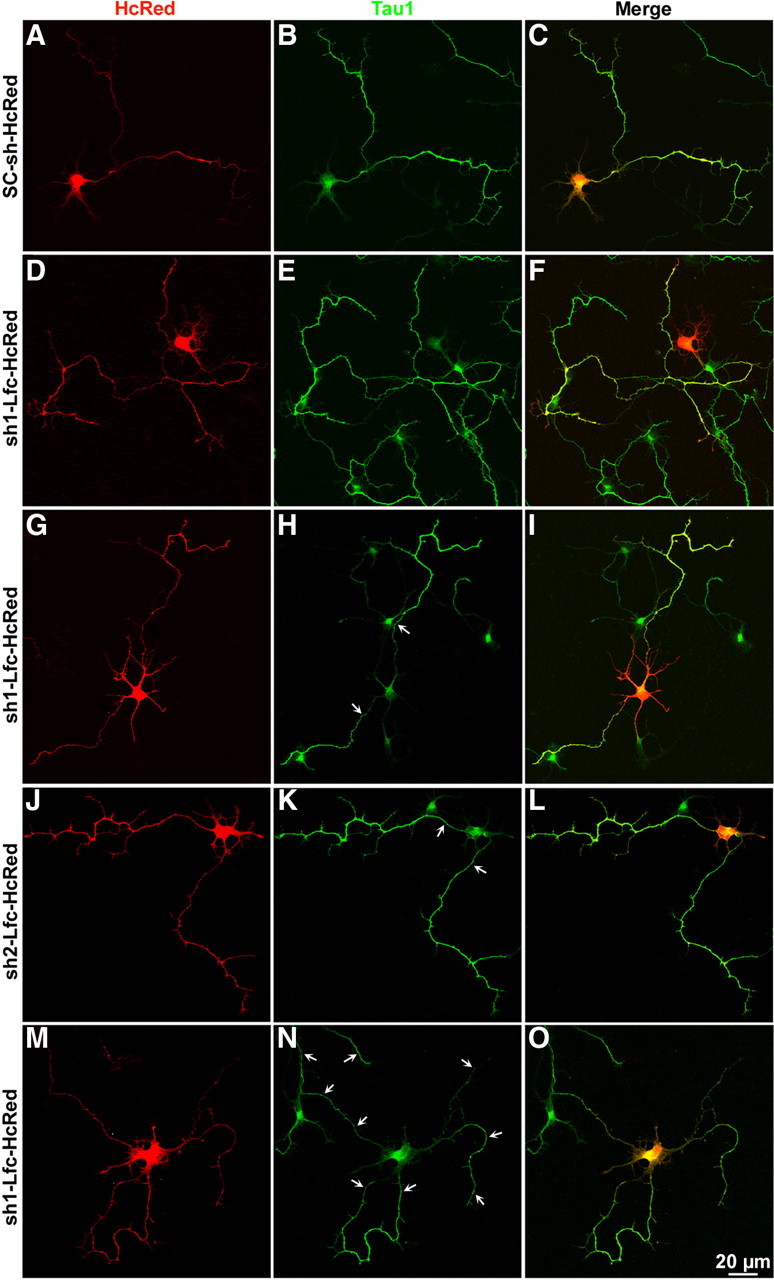
Lfc suppression enhances axon elongation and induces formation of supernumerary axons. A–O, Confocal images showing examples of neurons treated with control sh-RNA-HcRed (SC-sh-HcRed), sh1-Lfc-HcRed, or sh2-Lfc-HcRed. Cultures were transfected with the corresponding plasmids (4 μg DNA each) 48 h after plating and fixed 1 d later. The cultures were counterstained with the mAb Tau-1 (green). Note that Lfc suppression increases axonal length, collateral branching, and the formation of supernumerary axons (arrows).
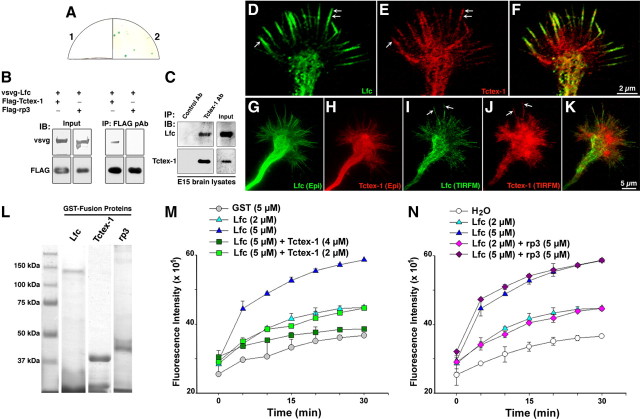
Tctex-1 interacts with Lfc and inhibits its GEF activity. A, Yeast two hybrid reveals an interaction between Tctex-1 and Lfc. 1, Negative control bait/prey, Lfc; 2, bait, Tctex-1/prey, Lfc. B, Immunoprecipitates of cell extracts from HEK cells transfected with vsvg-Lfc plus Flag-Tctex-1 or Flag-rp3. Note that Flag-Tctex-1, but not Flag-rp3, immunoprecipitated vsvg-Lfc. C, Control antibody (Control Ab) and Tctex-1 antibody (Tctex-1 Ab) immunoprecipitates performed on embryonic day 15 (E15) rat brain lysates and probed for Tctex-1 or Lfc; note the presence of Lfc in the Tctex-1 immunoprecipitates. D–F, A high-power confocal view of a large axonal growth cone from a stage 2–3 neuron showing the distribution of Lfc (green) and Tctex-1 (red). Note that both proteins colocalized within the growth cone C domain and at the distal end of filopodial extensions (arrows); Tctex-1 immunofluorescence was also present at the transition zone between the C and P domains, as well as at the base of filopodial extensions. G–K, A series of images showing the distribution of Lfc (green) and Tctex-1 (red) in an axonal growth cone as revealed by epifluorescence and TIRFM. Note that TIRFM images also revealed colocalization of Lfc and Tctex-1 within the growth cone P domain, including filopodial extensions (arrows). L, Coomassie blue staining of GST-fusion proteins used for the Lfc GEF assay. M, Graphs showing the GEF activity of GST-Lfc in the presence or absence of GST-Tctex-1. Note that GST-Lfc activity is inhibited in a dose-dependent manner by GST-Tctex-1; this inhibition was statistically significant (p < 0.001) after 5 min of initiating the assay. N, A similar GEF assay showing the GST-rp3 had no effect on GST-Lfc activity. IP, Immunoprecipitates; IB, immunoblots; pAb, polyclonal antibody.
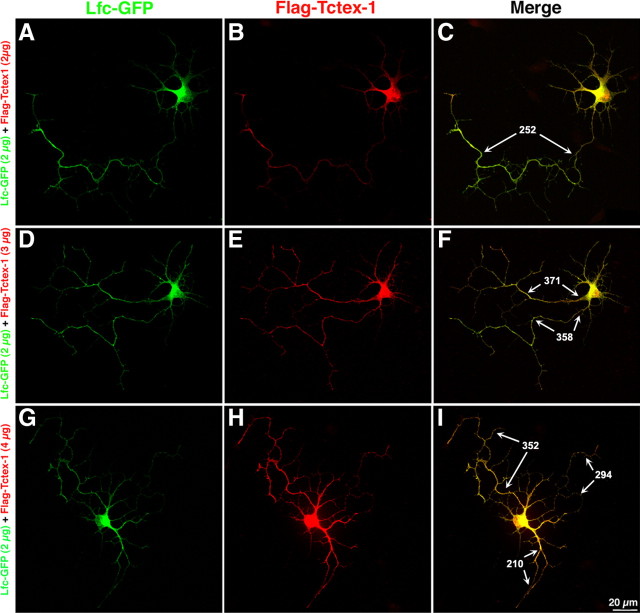
Tctex-1 counteracts the inhibitory effect of Lfc on axon formation. A–I, Confocal images showing hippocampal pyramidal neurons after cotransfection of Lfc-GFP (green) and Flag-Tctex-1 (red). Neurons were transfected 4 h after plating and fixed 20 h later. Note that ectopic expression of Tctex-1 promotes axon formation, elongation, and branching in neurons coexpressing Lfc and that this effect was dose dependent. The numbers in C, F, and I correspond to the length (micrometers) of axonal processes.
Similar articles
-
Distinct roles for the two Rho GDP/GTP exchange factor domains of kalirin in regulation of neurite growth and neuronal morphology.J Neurosci. 2001 Nov 1;21(21):8426-34. doi: 10.1523/JNEUROSCI.21-21-08426.2001. J Neurosci. 2001. PMID: 11606631 Free PMC article.
-
A novel FERM domain including guanine nucleotide exchange factor is involved in Rac signaling and regulates neurite remodeling.J Neurosci. 2002 Oct 1;22(19):8504-13. doi: 10.1523/JNEUROSCI.22-19-08504.2002. J Neurosci. 2002. PMID: 12351724 Free PMC article.
-
Golgi-resident TRIO regulates membrane trafficking during neurite outgrowth.J Biol Chem. 2019 Jul 12;294(28):10954-10968. doi: 10.1074/jbc.RA118.007318. Epub 2019 May 31. J Biol Chem. 2019. PMID: 31152060 Free PMC article.
-
Involvement of p114-RhoGEF and Lfc in Wnt-3a- and dishevelled-induced RhoA activation and neurite retraction in N1E-115 mouse neuroblastoma cells.Mol Biol Cell. 2010 Oct 15;21(20):3590-600. doi: 10.1091/mbc.E10-02-0095. Epub 2010 Sep 1. Mol Biol Cell. 2010. PMID: 20810787 Free PMC article.
-
The guanine nucleotide exchange factor Tiam1: a Janus-faced molecule in cellular signaling.Cell Signal. 2014 Mar;26(3):483-91. doi: 10.1016/j.cellsig.2013.11.034. Epub 2013 Dec 2. Cell Signal. 2014. PMID: 24308970 Review.
Cited by
-
Tctex-1, a novel interaction partner of Kidney Injury Molecule-1, is required for efferocytosis.J Cell Physiol. 2018 Oct;233(10):6877-6895. doi: 10.1002/jcp.26578. Epub 2018 Apr 25. J Cell Physiol. 2018. PMID: 29693725 Free PMC article.
-
Neurotoxicity of the pesticide rotenone on neuronal polarization: a mechanistic approach.Neural Regen Res. 2019 May;14(5):762-766. doi: 10.4103/1673-5374.249847. Neural Regen Res. 2019. PMID: 30688258 Free PMC article. Review.
-
Regulation of spine density and morphology by IQGAP1 protein domains.PLoS One. 2013;8(2):e56574. doi: 10.1371/journal.pone.0056574. Epub 2013 Feb 18. PLoS One. 2013. PMID: 23441206 Free PMC article.
-
Neuronal Polarity: Positive and Negative Feedback Signals.Front Cell Dev Biol. 2019 Apr 24;7:69. doi: 10.3389/fcell.2019.00069. eCollection 2019. Front Cell Dev Biol. 2019. PMID: 31069225 Free PMC article.
-
Changes in the GEF-H1 pathways after traumatic brain injury.J Neurotrauma. 2013 Aug 15;30(16):1449-56. doi: 10.1089/neu.2012.2673. Epub 2013 Jul 30. J Neurotrauma. 2013. PMID: 23611588 Free PMC article.
References
-
- Aijaz S, D'Atri F, Citi S, Balda MS, Matter K. Binding of GEF-H1 to the tight junction-associated adaptor cingulin results in inhibition of Rho signaling and G1/S phase transition. Dev Cell. 2005;8:777–786. - PubMed
-
- Arimura A, Kaibuchi K. Neuronal polarity: from extracellular signals to intracellular mechanisms. Nat Rev Neurosci. 2007;8:194–205. - PubMed
-
- Bhupinder P, Vohra S, Fu M, Heuckeroth RO. Protein kinase Czeta and glycogen synthase kinase-3beta control neuronal polarity in developing rodent enteric neurons, whereas SMAD-specific E3 ubiquitin protein ligase 1 promotes neurite growth but does not influence polarity. J Neurosci. 2007;27:9458–9468. - PMC - PubMed
-
- Birkenfeld J, Nalbant P, Yoon S, Bokoch G. Cellular functions of GEF-H1, a microtubule regulated Rho-GEF: is altered GEF-H1 activity a crucial determinant of disease pathogenesis? Trends Cell Biol. 2008;18:210–219. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous