

Ash2l interacts with Tbx1 and is required during early embryogenesis
- PMID: 20463296
- PMCID: PMC3017365
- DOI: 10.1258/ebm.2010.009318
Ash2l interacts with Tbx1 and is required during early embryogenesis
Abstract
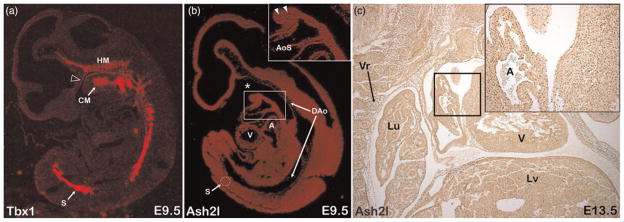
TBX1 encodes a DNA binding transcription factor that is commonly deleted in human DiGeorge syndrome and plays an important role in heart development. Mechanisms of Tbx1 function, such as Tbx1 interacting regulatory proteins and transcriptional target specificity, are largely unknown. Ash2l is the mammalian homolog of Drosophila Ash2 (absent small homeotic 2) and is a core component of a multimeric histone methyltransferase complex that epigenetically regulates transcription via methylation of histone lysine residues. We undertook an unbiased yeast two-hybrid screen to look for functionally relevant Tbx1-interacting proteins and report a physical and functional interaction between Tbx1 and Ash2l. Tbx1 interacts with Ash2l in both yeast and mammalian cells and Ash2l acts as a transcriptional co-activator in luciferase reporter assays. Expression analysis shows that Tbx1 and Ash2l have overlapping mRNA and protein expression patterns during development. By generating an Ash2l knockout mouse utilizing gene-trap technology, we show that although Ash2l heterozygous mice are normal, Ash2l-null embryos die early during gestation. Thus, Ash2l is required for the earliest stages of embryogenesis. Furthermore, our finding of a physical interaction between Tbx1 and Ash2l suggest that at least some functions of Tbx1 may be mediated by direct interactions with a histone methyltransferase complex.
Figures


Similar articles
-
ASH2L is involved in promotion of endometrial cancer progression via upregulation of PAX2 transcription.Cancer Sci. 2020 Jun;111(6):2062-2077. doi: 10.1111/cas.14413. Epub 2020 May 23. Cancer Sci. 2020. PMID: 32279431 Free PMC article.
-
Tbx1 regulates progenitor cell proliferation in the dental epithelium by modulating Pitx2 activation of p21.Dev Biol. 2010 Nov 15;347(2):289-300. doi: 10.1016/j.ydbio.2010.08.031. Epub 2010 Sep 15. Dev Biol. 2010. PMID: 20816801 Free PMC article.
-
Transcription factor Ap2delta associates with Ash2l and ALR, a trithorax family histone methyltransferase, to activate Hoxc8 transcription.Proc Natl Acad Sci U S A. 2008 May 27;105(21):7472-7. doi: 10.1073/pnas.0711896105. Epub 2008 May 21. Proc Natl Acad Sci U S A. 2008. PMID: 18495928 Free PMC article.
-
Tbx1: Transcriptional and Developmental Functions.Curr Top Dev Biol. 2017;122:223-243. doi: 10.1016/bs.ctdb.2016.08.002. Epub 2016 Sep 1. Curr Top Dev Biol. 2017. PMID: 28057265 Review.
-
T-Box Genes in Human Development and Disease.Curr Top Dev Biol. 2017;122:383-415. doi: 10.1016/bs.ctdb.2016.08.006. Epub 2016 Oct 5. Curr Top Dev Biol. 2017. PMID: 28057271 Review.
Cited by
-
Elucidating the mechanisms of transcription regulation during heart development by next-generation sequencing.J Hum Genet. 2016 Jan;61(1):5-12. doi: 10.1038/jhg.2015.84. Epub 2015 Jul 23. J Hum Genet. 2016. PMID: 26202577 Review.
-
Absent, Small, or Homeotic 2-Like-Mediated H3K4 Methylation and Nephrogenesis.J Am Soc Nephrol. 2025 May 1;36(5):798-811. doi: 10.1681/ASN.0000000600. Epub 2025 Jan 7. J Am Soc Nephrol. 2025. PMID: 39774048
-
Chromatin remodeling and bivalent histone modifications in embryonic stem cells.EMBO Rep. 2015 Dec;16(12):1609-19. doi: 10.15252/embr.201541011. Epub 2015 Nov 9. EMBO Rep. 2015. PMID: 26553936 Free PMC article. Review.
-
Epigenetic regulation of cardiac development and function by polycomb group and trithorax group proteins.Dev Dyn. 2012 Jun;241(6):1021-33. doi: 10.1002/dvdy.23796. Epub 2012 May 8. Dev Dyn. 2012. PMID: 22514007 Free PMC article. Review.
-
Sequential deregulation of histone marks, chromatin accessibility and gene expression in response to PROTAC-induced degradation of ASH2L.Sci Rep. 2023 Dec 19;13(1):22565. doi: 10.1038/s41598-023-49284-x. Sci Rep. 2023. PMID: 38114530 Free PMC article.
References
-
- Yagi H, Furutani Y, Hamada H, Sasaki T, Asakawa S, Minoshima S, Ichida F, Joo K, Kimura M, Imamura S, Kamatani N, Momma K, Takao A, Nakazawa M, Shimizu N, Matsuoka R. Role of TBX1 in human del22q11.2 syndrome. Lancet. 2003;362:1366–73. - PubMed
-
- Stennard FA, Costa MW, Elliott DA, Rankin S, Haast SJ, Lai D, McDonald LP, Niederreither K, Dolle P, Bruneau BG, Zorn AM, Harvey RP. Cardiac T-box factor Tbx20 directly interacts with Nkx2-5, GATA4, and GATA5 in regulation of gene expression in the developing heart. Dev Biol. 2003;262:206–24. - PubMed
-
- Farin HF, Bussen M, Schmidt MK, Singh MK, Schuster-Gossler K, Kispert A. Transcriptional repression by the T-box proteins Tbx18 and Tbx15 depends on Groucho corepressors. J Biol Chem. 2007;282:25748–59. - PubMed
-
- Hiroi Y, Kudoh S, Monzen K, Ikeda Y, Yazaki Y, Nagai R, Komuro I. Tbx5 associates with Nkx2–5 and synergistically promotes cardiomyocyte differentiation. Nat Genet. 2001;28:276–80. - PubMed
-
- Farin HF, Mansouri A, Petry M, Kispert A. T-box protein Tbx18 interacts with the paired box protein Pax3 in the development of the paraxial mesoderm. J Biol Chem. 2008;283:25372–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases