

Mitochondrial iron metabolism and its role in neurodegeneration
- PMID: 20463401
- PMCID: PMC3085540
- DOI: 10.3233/JAD-2010-100354
Mitochondrial iron metabolism and its role in neurodegeneration
Abstract
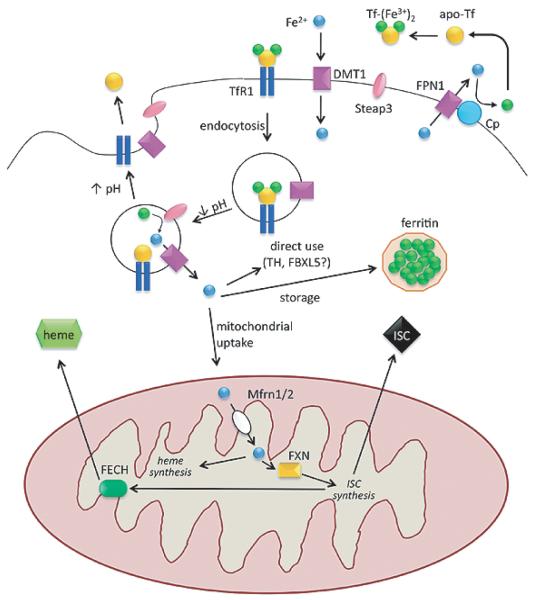
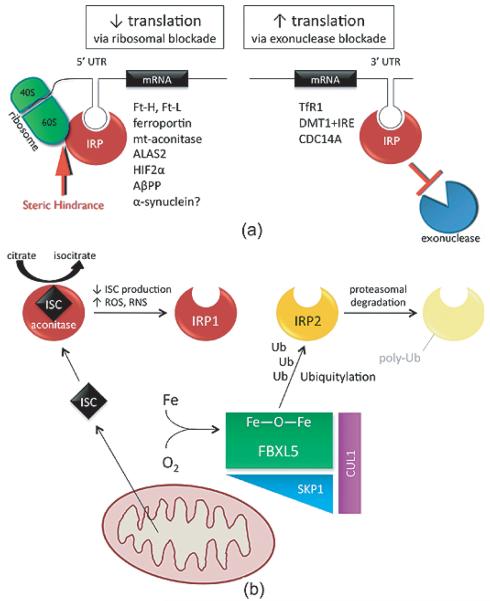
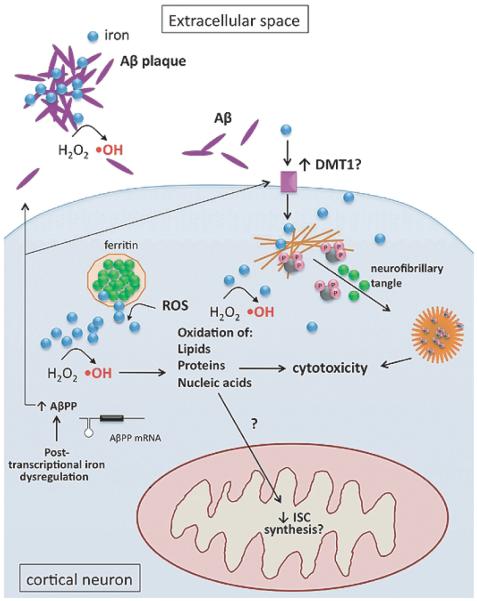
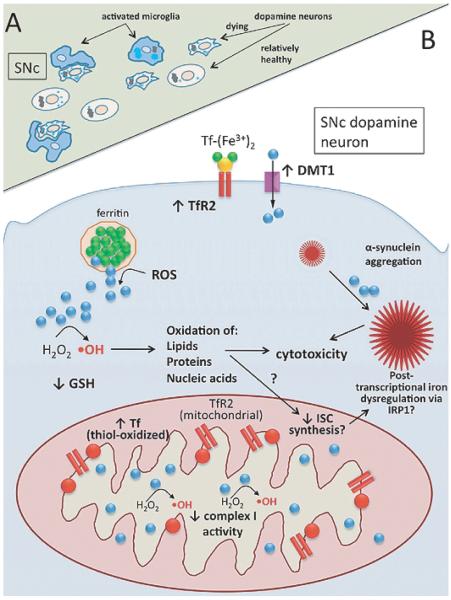
In addition to their well-established role in providing the cell with ATP, mitochondria are the source of iron-sulfur clusters (ISCs) and heme - prosthetic groups that are utilized by proteins throughout the cell in various critical processes. The post-transcriptional system that mammalian cells use to regulate intracellular iron homeostasis depends, in part, upon the synthesis of ISCs in mitochondria. Thus, proper mitochondrial function is crucial to cellular iron homeostasis. Many neurodegenerative diseases are marked by mitochondrial impairment, brain iron accumulation, and oxidative stress - pathologies that are inter-related. This review discusses the physiological role that mitochondria play in cellular iron homeostasis and, in so doing, attempts to clarify how mitochondrial dysfunction may initiate and/or contribute to iron dysregulation in the context of neurodegenerative disease. We review what is currently known about the entry of iron into mitochondria, the ways in which iron is utilized therein, and how mitochondria are integrated into the system of iron homeostasis in mammalian cells. Lastly, we turn to recent advances in our understanding of iron dysregulation in two neurodegenerative diseases (Alzheimer's disease and Parkinson's disease), and discuss the use of iron chelation as a potential therapeutic approach to neurodegenerative disease.
Figures
References
-
- Herrero-Mendez A, Almeida A, Fernandez E, Maestre C, Moncada S, Bolanos JP. The bioenergetic and antioxidant status of neurons is controlled by continuous degradation of a key glycolytic enzyme by APC/C-Cdh1. Nat Cell Biol. 2009;11:747–752. - PubMed
-
- Sheftel A, Stehling O, Lill R. Iron-sulfur proteins in health and disease. Trends Endocrinol Metab. 2010;21:302–314. - PubMed
-
- Furuyama K, Kaneko K, Vargas PD. Heme as a magnificent molecule with multiple missions: heme determines its own fate and governs cellular homeostasis. Tohoku J Exp Med. 2007;213:1–16. - PubMed
-
- Garrick MD, Garrick LM. Cellular iron transport. Biochim Biophys Acta. 2009;1790:309–325. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
