

The gene transformer-2 of Anastrepha fruit flies (Diptera, Tephritidae) and its evolution in insects
- PMID: 20465812
- PMCID: PMC2885393
- DOI: 10.1186/1471-2148-10-140
The gene transformer-2 of Anastrepha fruit flies (Diptera, Tephritidae) and its evolution in insects
Abstract
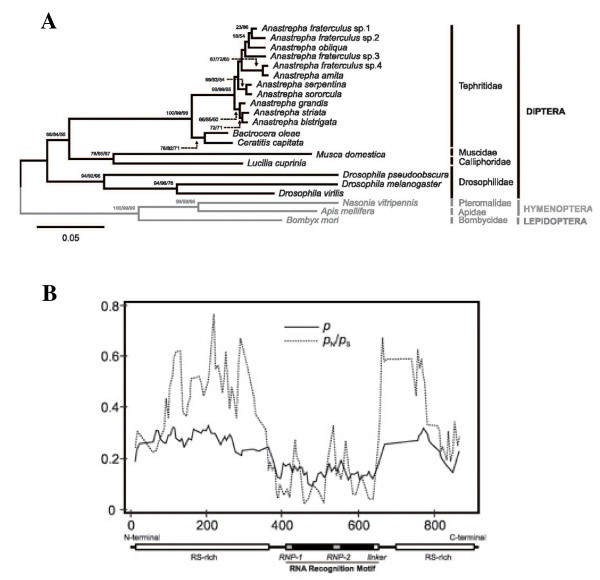
Background: In the tephritids Ceratitis, Bactrocera and Anastrepha, the gene transformer provides the memory device for sex determination via its auto-regulation; only in females is functional Tra protein produced. To date, the isolation and characterisation of the gene transformer-2 in the tephritids has only been undertaken in Ceratitis, and it has been shown that its function is required for the female-specific splicing of doublesex and transformer pre-mRNA. It therefore participates in transformer auto-regulatory function. In this work, the characterisation of this gene in eleven tephritid species belonging to the less extensively analysed genus Anastrepha was undertaken in order to throw light on the evolution of transformer-2.
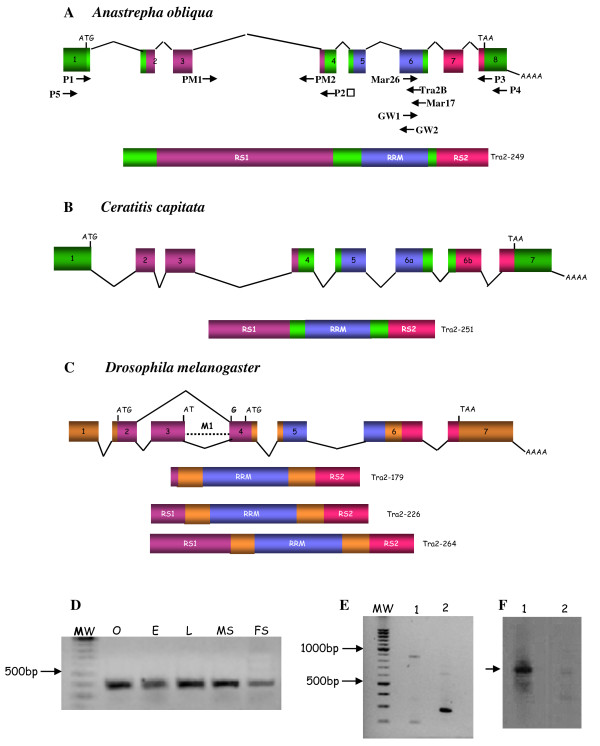
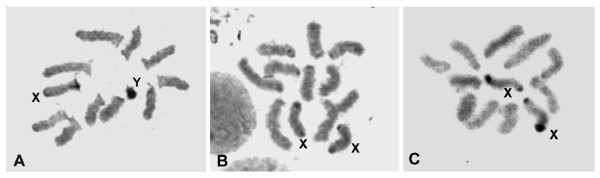
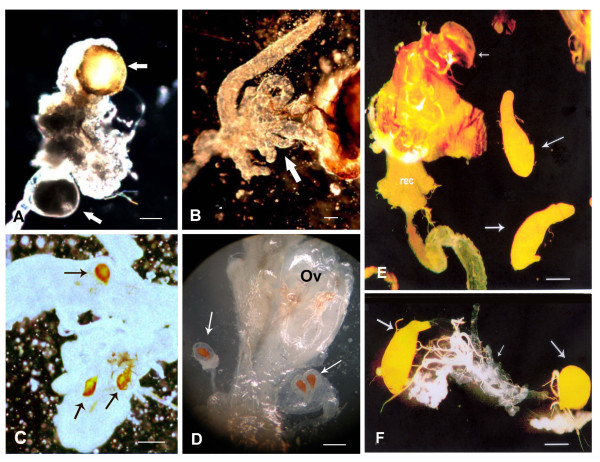
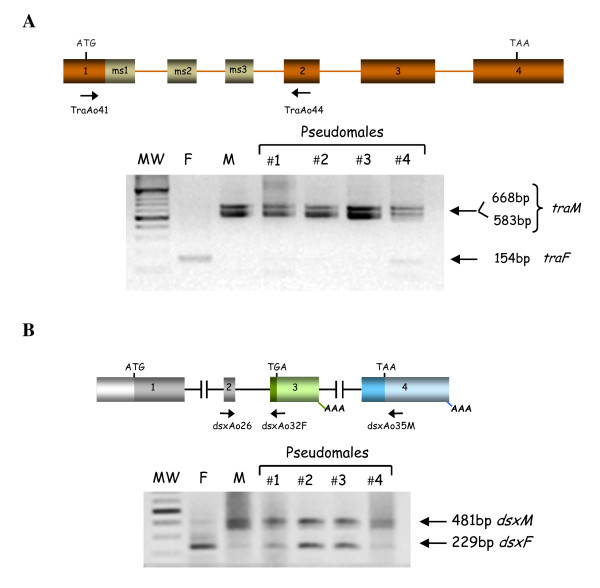
Results: The gene transformer-2 produces a protein of 249 amino acids in both sexes, which shows the features of the SR protein family. No significant partially spliced mRNA isoform specific to the male germ line was detected, unlike in Drosophila. It is transcribed in both sexes during development and in adult life, in both the soma and germ line. The injection of Anastrepha transformer-2 dsRNA into Anastrepha embryos caused a change in the splicing pattern of the endogenous transformer and doublesex pre-mRNA of XX females from the female to the male mode. Consequently, these XX females were transformed into pseudomales. The comparison of the eleven Anastrepha Transformer-2 proteins among themselves, and with the Transformer-2 proteins of other insects, suggests the existence of negative selection acting at the protein level to maintain Transformer-2 structural features.
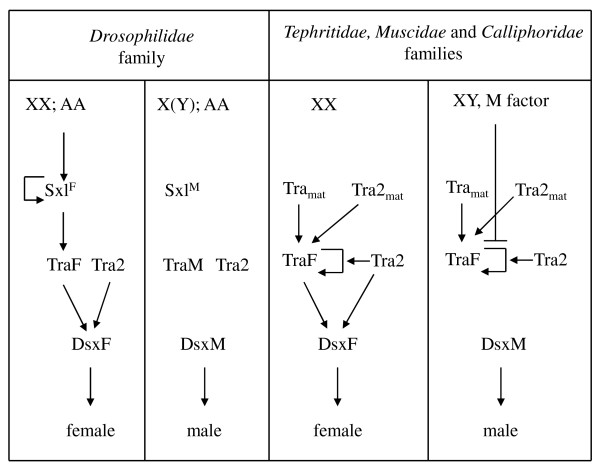
Conclusions: These results indicate that transformer-2 is required for sex determination in Anastrepha through its participation in the female-specific splicing of transformer and doublesex pre-mRNAs. It is therefore needed for the auto-regulation of the gene transformer. Thus, the transformer/transfomer-2 > doublesex elements at the bottom of the cascade, and their relationships, probably represent the ancestral state (which still exists in the Tephritidae, Calliphoridae and Muscidae lineages) of the extant cascade found in the Drosophilidae lineage (in which tra is just another component of the sex determination gene cascade regulated by Sex-lethal). In the phylogenetic lineage that gave rise to the drosophilids, evolution co-opted for Sex-lethal, modified it, and converted it into the key gene controlling sex determination.
Figures
References
-
- Sánchez L, Gorfinkiel N, Guerrero I. In: Comprehensive Molecular Insect Science. Gilbert LI, Iatrou K, Gill SS, editor. Vol. 1. Oxford: Elsevier Pergamon; 2005. Sex determination and the development of the genital disc; pp. 1–38. full_text.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
