

Activation of alpha1 and alpha2 noradrenergic receptors exert opposing effects on excitability of main olfactory bulb granule cells
- PMID: 20466037
- PMCID: PMC2904409
- DOI: 10.1016/j.neuroscience.2010.05.010
Activation of alpha1 and alpha2 noradrenergic receptors exert opposing effects on excitability of main olfactory bulb granule cells
Abstract
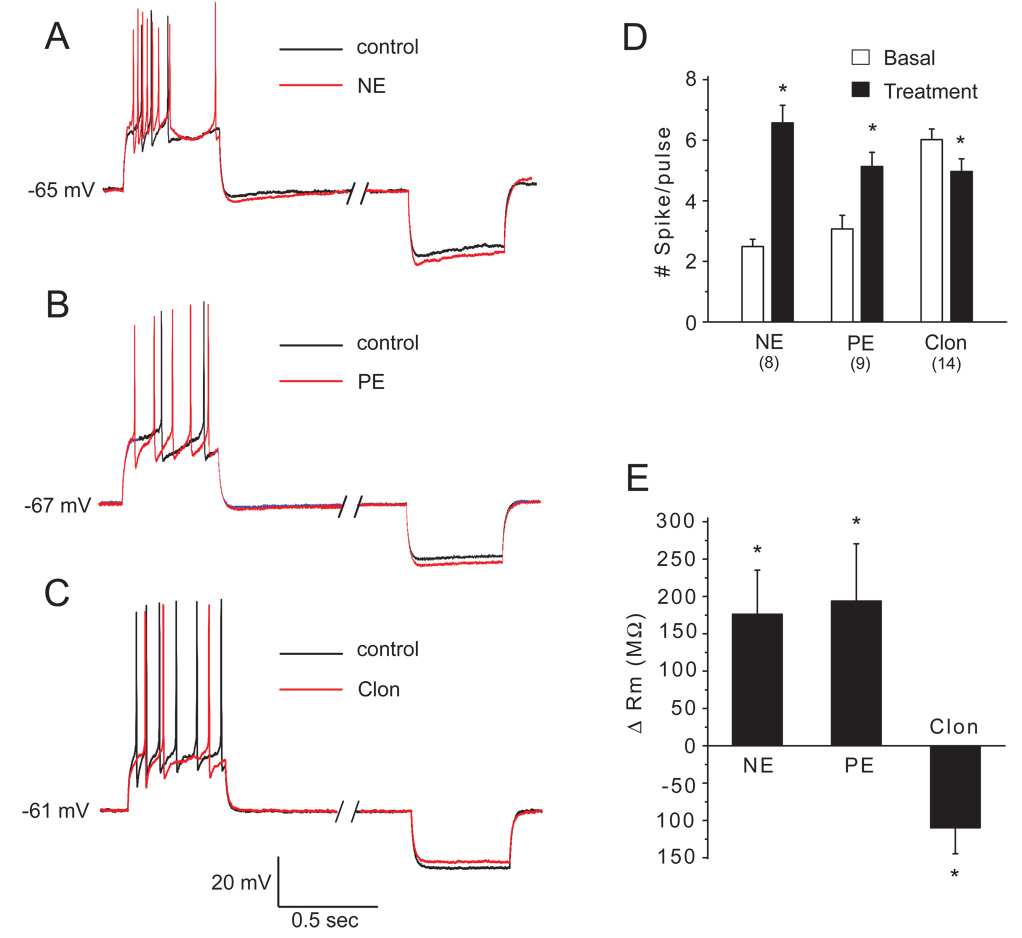
The mammalian main olfactory bulb (MOB) receives a dense noradrenergic innervation from the pontine nucleus locus coeruleus that is important for neonatal odor preference learning and odor processing in mature animals. Modulation of GABAergic granule cells (GCs) is thought to play a key role in the net functional impact of norepinephrine (NE) release in the MOB, yet there are few direct studies of the influence of NE on these cells. In the present study we investigated noradrenergic modulation of GC excitability using electrophysiological approaches in rat MOB slices. A moderate concentration of NE (10 microM) and the alpha1 receptor agonist phenylephrine (10 microM) depolarized and increased spontaneous or current injection-evoked spiking in GCs. By contrast, low NE concentrations (0.1-1.0 microM) or the alpha2 receptor agonist clonidine (Clon, 10 microM) hyperpolarized and decreased the discharge of GCs. The effects of NE (10 microM) were blocked by antagonism of alpha1 and alpha2 receptors. Inhibitory effects of low NE concentrations were blocked or converted to excitatory responses by alpha2 receptor blockade, whereas excitatory effects of the moderate NE concentration were converted to inhibitory responses after alpha1 receptor blockade. NE (10 microM) and phenylephrine elicited inward currents that reversed near the potassium equilibrium potential. The effects of NE and phenylephrine were associated with increased membrane input resistance. Clonidine elicited an outward current associated with decreased membrane input resistance that reversed near the potassium equilibrium potential. These results indicate that alpha1 and alpha2 receptor activation exert opposing effects on GC excitability. Low concentrations of NE acting via alpha2 receptors suppress GC excitability, while higher concentrations of NE acting at alpha1 receptors increase GC excitability. These findings are consistent with recent findings that alpha1 and alpha2 receptor activation increase and decrease, respectively, GABAergic inhibition of mitral cells. The differential affinities of alpha1 and alpha2 noradrenergic receptor subtypes may allow for differential modulation of GABA release and olfactory processing as a function of the level of NE release, which in turn, is regulated by behavioral state.
Copyright (c) 2010 IBRO. Published by Elsevier Ltd. All rights reserved.
Figures




References
-
- Berridge C, Waterhouse B. The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain Res Rev. 2003;42:33–84. - PubMed
-
- Czesnik D, Nezlin L, Rabba J, Muller B, Schild D. Noradrenergic modulation of calcium currents and synaptic transmission in the olfactory bulb of Xenopus laevis. Eur J Neursci. 2001;13:1093–1100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
