

IL-27 inhibits OSM-mediated TNF-alpha and iNOS gene expression in microglia
- PMID: 20468050
- PMCID: PMC3378052
- DOI: 10.1002/glia.20989
IL-27 inhibits OSM-mediated TNF-alpha and iNOS gene expression in microglia
Abstract
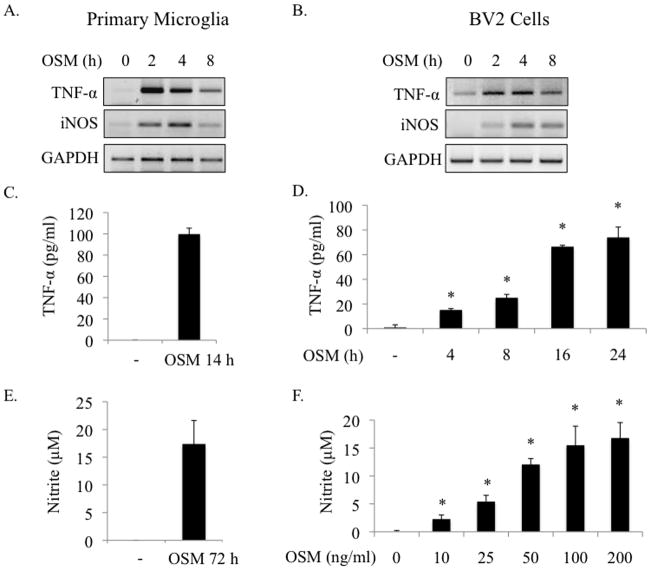
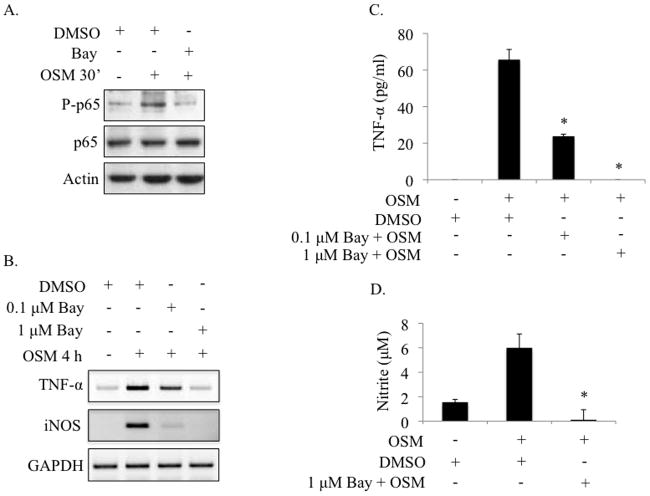
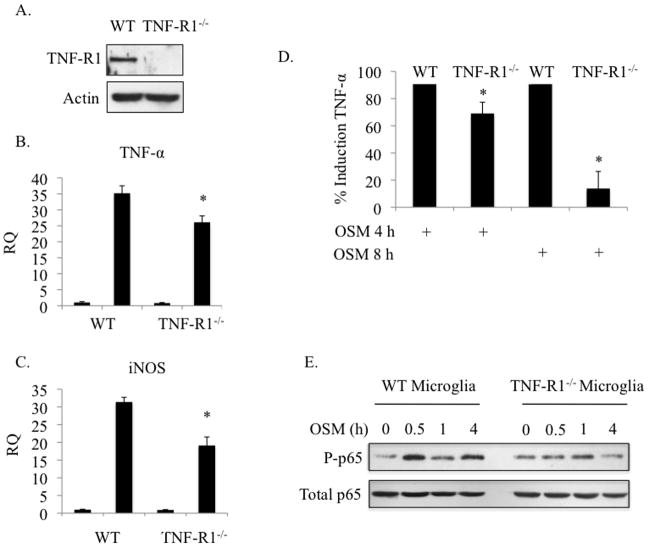
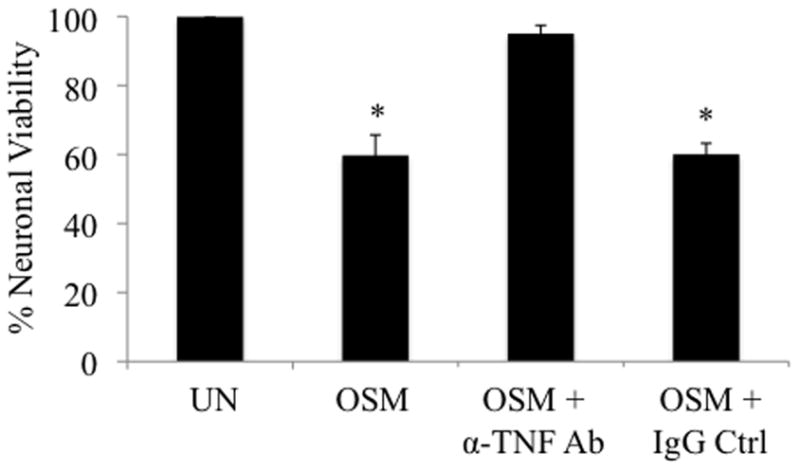
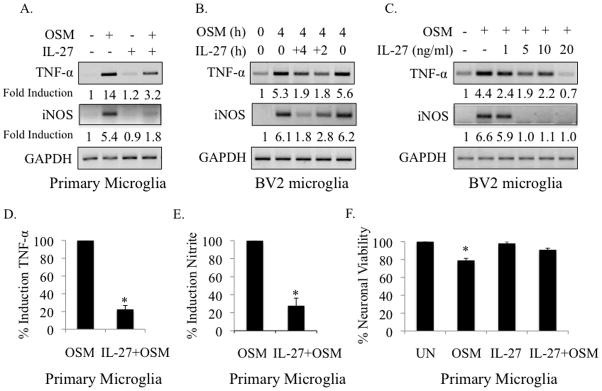
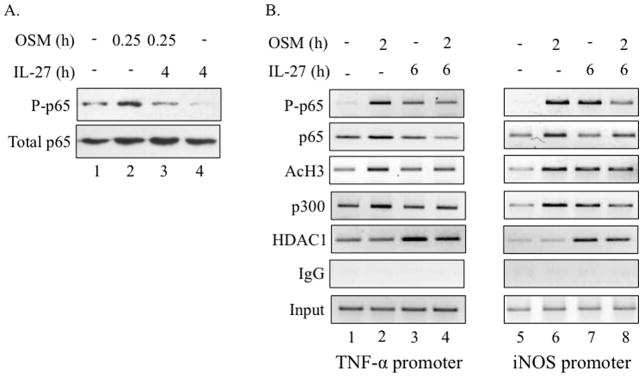
Elevated levels of Oncostatin M (OSM), an interleukin-6 family cytokine, have been observed in multiple sclerosis (MS), HIV-associated neurocognitive disorder (HAND), and glioblastoma (GBM); however, its effects within the CNS are not well understood. OSM regulates gene expression primarily by activating the JAK/STAT, NF-kappaB, and/or MAPK pathways, in a cell-type specific manner. In our studies, OSM induces the production of the proinflammatory cytokine tumor necrosis factor-alpha (TNF-alpha) and inducible nitric oxide synthase (iNOS) from microglia in an NF-kappaB-dependent manner. This expression also partially requires the intermediate production of TNF-alpha and subsequent NF-kappaB activation via TNF-R1. We also demonstrate that OSM-induced TNF-alpha production from microglia is neurotoxic. The IL-12 family member, IL-27, suppresses OSM-mediated TNF-alpha and iNOS expression at the transcriptional level by inhibiting activation of the NF-kappaB pathway, and rescues the neurotoxicity induced by OSM-stimulated microglia. These studies are the first to demonstrate the proinflammatory effects of OSM in microglia, and also identify IL-27 as a novel inhibitor of inflammatory processes in these cells.
Copyright 2010 Wiley-Liss, Inc.
Figures


References
-
- Aggarwal BB. Signalling pathways of the TNF superfamily: a double-edged sword. Nat Rev Immunol. 2003;3(9):745–56. - PubMed
-
- Allan SM, Rothwell NJ. Cytokines and acute neurodegeneration. Nat Rev Neurosci. 2001;2(10):734–44. - PubMed
-
- Arnett HA, Mason J, Marino M, Suzuki K, Matsushima GK, Ting JP. TNF alpha promotes proliferation of oligodendrocyte progenitors and remyelination. Nat Neurosci. 2001;4(11):1116–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
