

Protein kinase C epsilon regulation of translocator protein (18 kDa) Tspo gene expression is mediated through a MAPK pathway targeting STAT3 and c-Jun transcription factors
- PMID: 20469933
- PMCID: PMC2902160
- DOI: 10.1021/bi100020e
Protein kinase C epsilon regulation of translocator protein (18 kDa) Tspo gene expression is mediated through a MAPK pathway targeting STAT3 and c-Jun transcription factors
Abstract
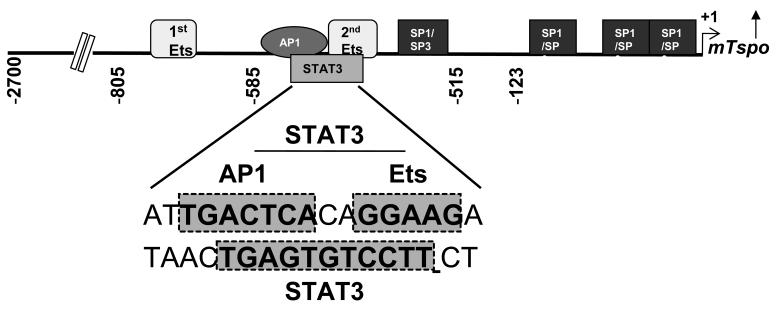
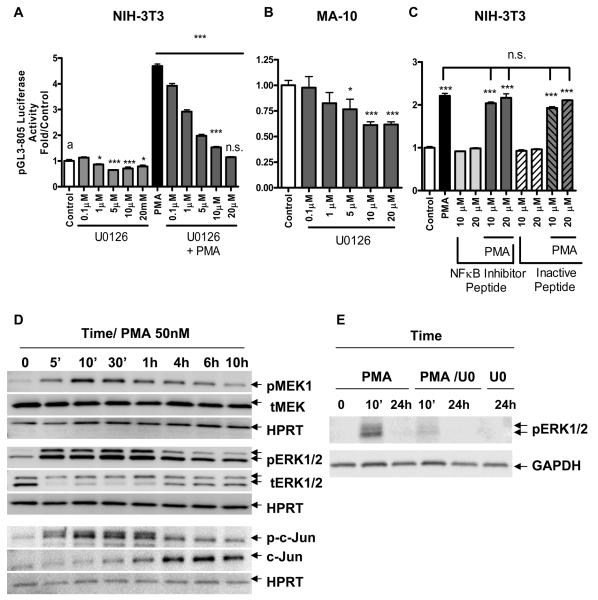
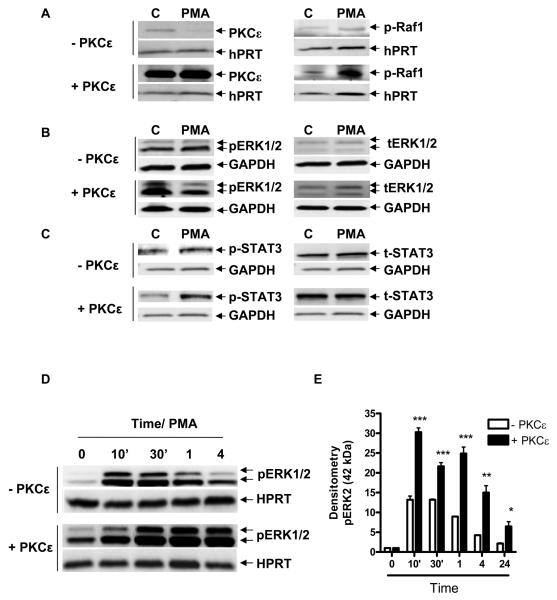
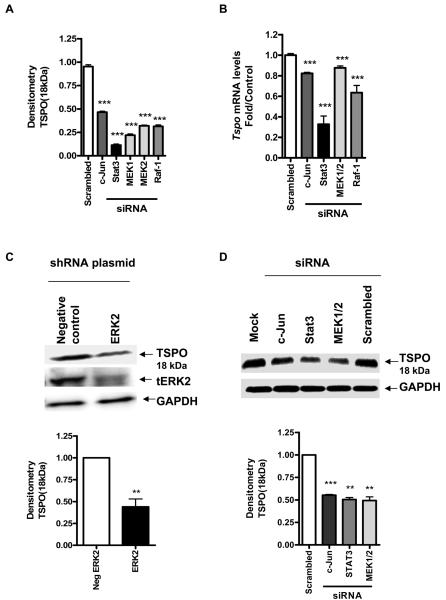
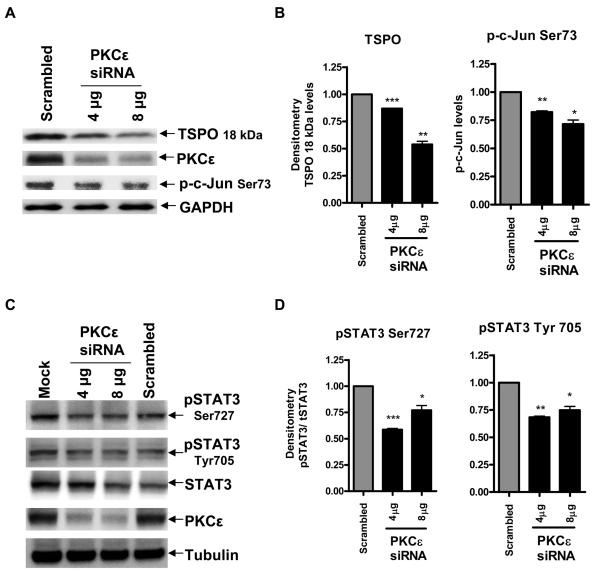
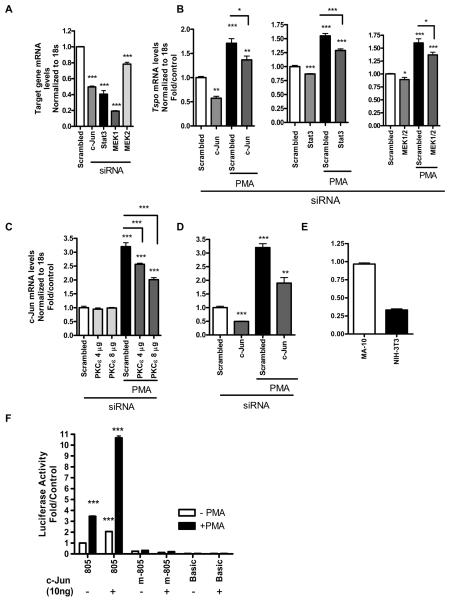
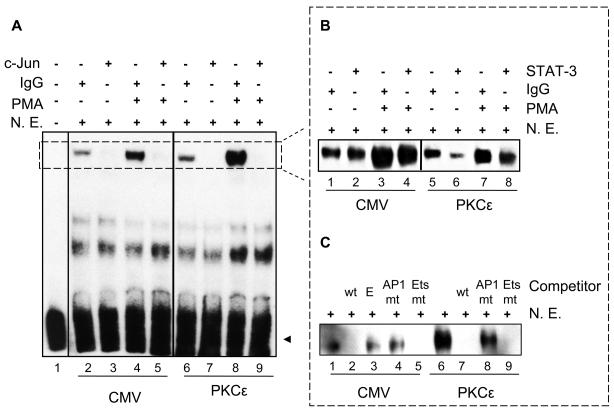
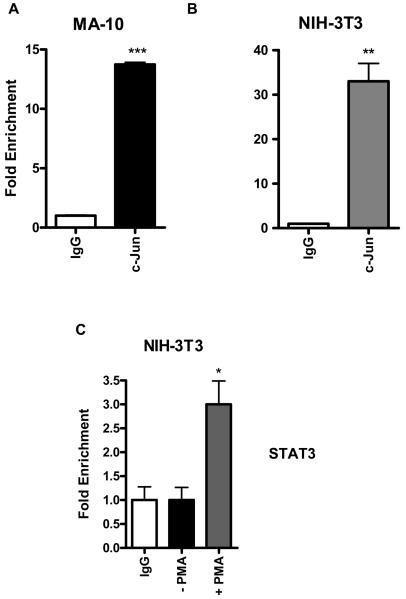
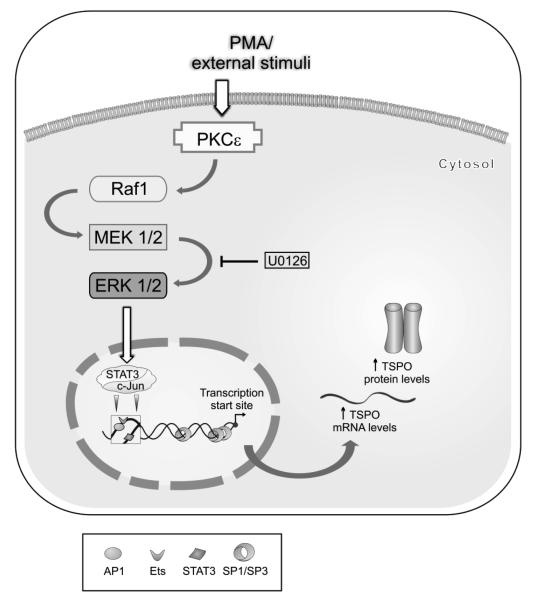
Translocator protein TSPO is an 18 kDa protein implicated in numerous cell functions and is highly expressed in secretory and glandular tissues, especially in steroidogenic cells. TSPO expression is altered in pathological conditions such as certain cancers and neurological diseases. In search of the factors regulating Tspo expression, we recently showed that high levels of TSPO in steroidogenic cells may be due to high constitutive expression of protein kinase Cepsilon (PKCepsilon), while phorbol 12-myristate 13-acetate (PMA) activation of PKCepsilon drives inducible TSPO expression in nonsteroidogenic cells, likely through activator protein 1 (AP1). In this study, we aimed to identify the signal transduction pathway through which PKCepsilon regulates Tspo gene expression. The MEK1/2 specific inhibitor U0126, but not NFkappaB inhibitors, reduced basal Tspo promoter activity in TSPO-rich steroidogenic cells (MA-10 Leydig), as well as basal and PMA-induced Tspo promoter levels in TSPO-poor nonsteroidogenic cells (NIH-3T3 fibroblasts). AP1 and signal transducer and activation of transcription 3 (STAT3) have binding sites in the Tspo promoter and are downstream targets of PKCepsilon and MAPK (Raf-1-ERK1/2) pathways. PKCepsilon overexpression induced STAT3 phosphorylation in NIH-3T3 cells, while PKCepsilon knockdown reduced STAT3 and c-Jun phosphorylation in Leydig cells. MEK1/2, ERK2, c-Jun, and STAT3 knockdown reduced Tspo mRNA and protein levels in Leydig cells. Additionally, Raf-1 reduced Tspo mRNA levels in the same cells. MEK1/2, c-Jun, and STAT3 knockdown also reduced basal as well as PMA-induced Tspo mRNA levels in NIH-3T3 cells. Together, these results demonstrate that PKCepsilon regulates Tspo gene expression through a MAPK (Raf-1-MEK1/2-ERK1/2) signal transduction pathway, acting at least in part through c-Jun and STAT3 transcription factors.
Figures
References
-
- Casellas P, Galiegue S, Basile AS. Peripheral benzodiazepine receptors and mitochondrial function. Neurochem. Int. 2002;40:475–486. - PubMed
-
- Papadopoulos V, Baraldi M, Guilarte TR, Knudsen TB, Lacapere JJ, Lindemann P, Norenberg MD, Nutt D, Weizman A, Zhang MR, Gavish M. Translocator protein (18kDa): new nomenclature for the peripheral-type benzodiazepine receptor based on its structure and molecular function. Trends Pharmacol. Sci. 2006;27:402–409. - PubMed
-
- Gavish M, Bachman I, Shoukrun R, Katz Y, Veenman L, Weisinger G, Weizman A. Enigma of the peripheral benzodiazepine receptor. Pharmacol. Rev. 1999;51:629–650. - PubMed
-
- Giatzakis C, Papadopoulos V. Differential utilization of the promoter of peripheral-type benzodiazepine receptor by steroidogenic versus nonsteroidogenic cell lines and the role of Sp1 and Sp3 in the regulation of basal activity. Endocrinology. 2004;145:1113–1123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
