

Multisensory integration: resolving sensory ambiguities to build novel representations
- PMID: 20471245
- PMCID: PMC2901182
- DOI: 10.1016/j.conb.2010.04.009
Multisensory integration: resolving sensory ambiguities to build novel representations
Abstract
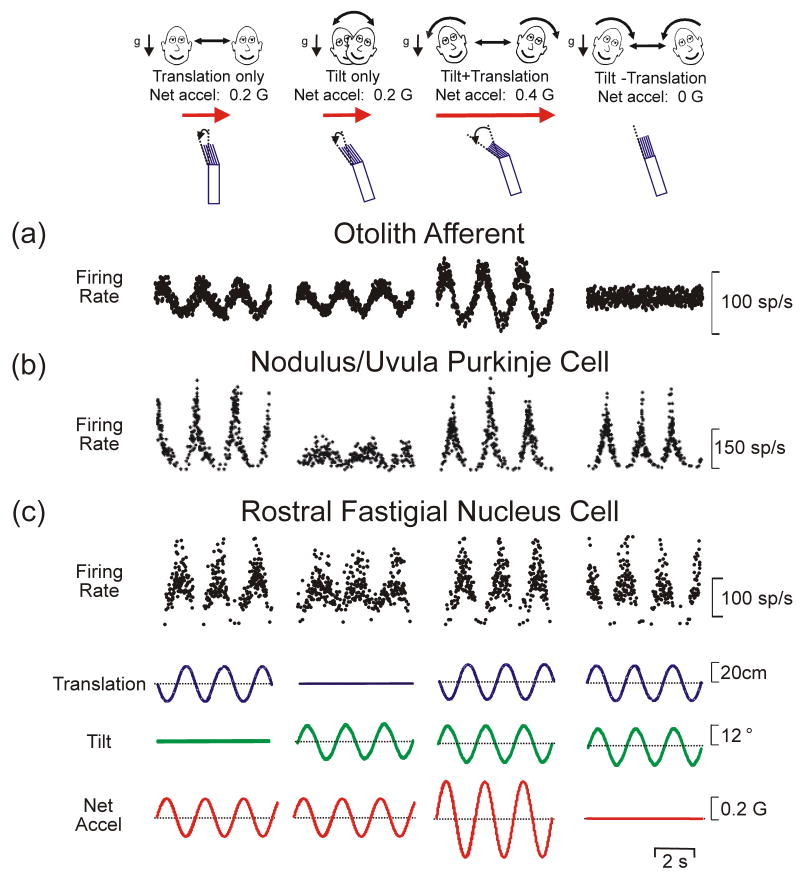
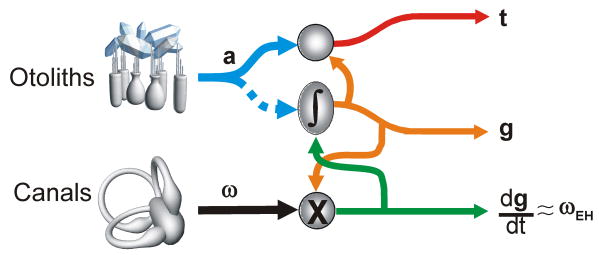
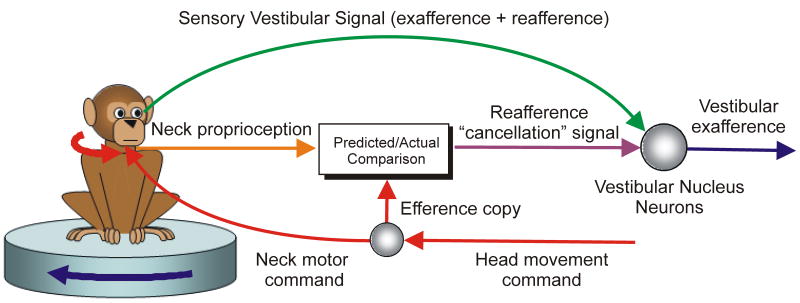
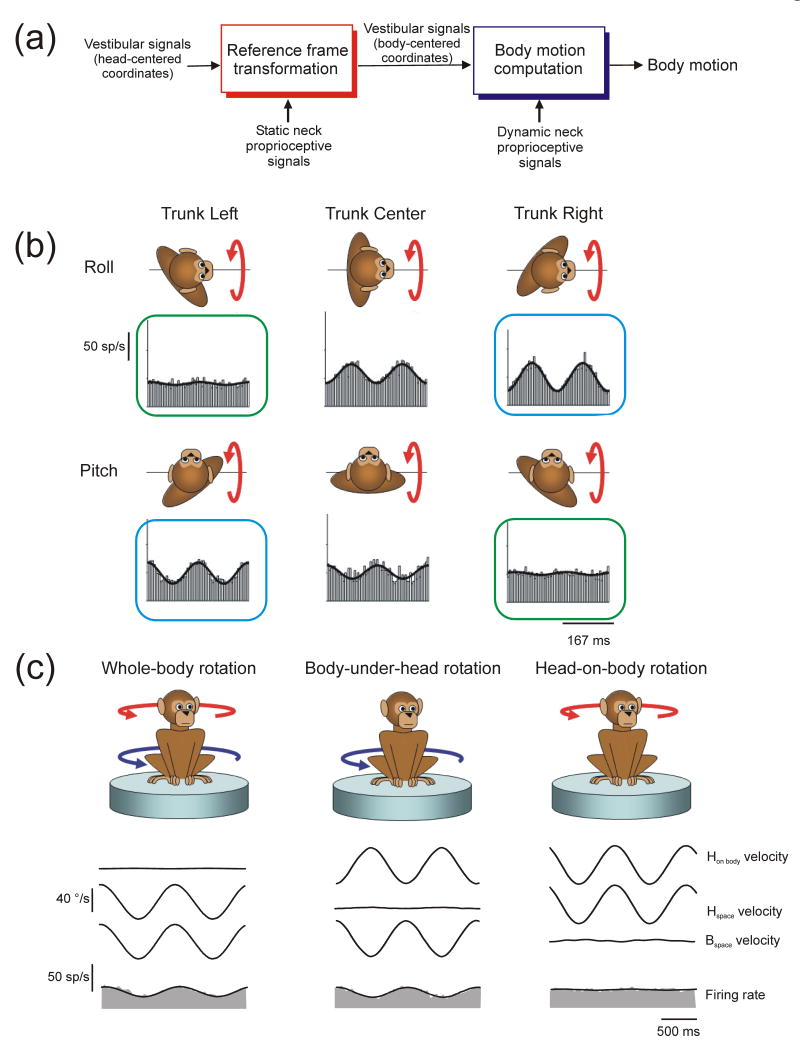
Multisensory integration plays several important roles in the nervous system. One is to combine information from multiple complementary cues to improve stimulus detection and discrimination. Another is to resolve peripheral sensory ambiguities and create novel internal representations that do not exist at the level of individual sensors. Here we focus on how ambiguities inherent in vestibular, proprioceptive and visual signals are resolved to create behaviorally useful internal estimates of our self-motion. We review recent studies that have shed new light on the nature of these estimates and how multiple, but individually ambiguous, sensory signals are processed and combined to compute them. We emphasize the need to combine experiments with theoretical insights to understand the transformations that are being performed.
2010 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
The internal representation of head orientation differs for conscious perception and balance control.J Physiol. 2017 Apr 15;595(8):2731-2749. doi: 10.1113/JP272998. Epub 2017 Feb 1. J Physiol. 2017. PMID: 28035656 Free PMC article.
-
Visual-vestibular cue integration for heading perception: applications of optimal cue integration theory.Eur J Neurosci. 2010 May;31(10):1721-9. doi: 10.1111/j.1460-9568.2010.07207.x. Eur J Neurosci. 2010. PMID: 20584175 Free PMC article. Review.
-
Fusion of vestibular and podokinesthetic information during self-turning towards instructed targets.Exp Brain Res. 2002 Jun;144(4):458-74. doi: 10.1007/s00221-002-1053-5. Epub 2002 Apr 25. Exp Brain Res. 2002. PMID: 12037631
-
Multisensory integration in the estimation of relative path length.Exp Brain Res. 2004 Jan;154(2):246-54. doi: 10.1007/s00221-003-1652-9. Epub 2003 Sep 6. Exp Brain Res. 2004. PMID: 14685814
-
The sense of self-motion, orientation and balance explored by vestibular stimulation.J Physiol. 2011 Feb 15;589(Pt 4):807-13. doi: 10.1113/jphysiol.2010.197665. Epub 2010 Oct 4. J Physiol. 2011. PMID: 20921198 Free PMC article. Review.
Cited by
-
Asymmetric multisensory interactions of visual and somatosensory responses in a region of the rat parietal cortex.PLoS One. 2013 May 7;8(5):e63631. doi: 10.1371/journal.pone.0063631. Print 2013. PLoS One. 2013. PMID: 23667650 Free PMC article.
-
The Motor Network Reduces Multisensory Illusory Perception.J Neurosci. 2018 Nov 7;38(45):9679-9688. doi: 10.1523/JNEUROSCI.3650-17.2018. Epub 2018 Sep 24. J Neurosci. 2018. PMID: 30249803 Free PMC article.
-
Metacognitive awareness in the sound-induced flash illusion.Philos Trans R Soc Lond B Biol Sci. 2023 Sep 25;378(1886):20220347. doi: 10.1098/rstb.2022.0347. Epub 2023 Aug 7. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 37545312 Free PMC article.
-
Simple spike dynamics of Purkinje cells in the macaque vestibulo-cerebellum during passive whole-body self-motion.Proc Natl Acad Sci U S A. 2020 Feb 11;117(6):3232-3238. doi: 10.1073/pnas.1915873117. Epub 2020 Jan 27. Proc Natl Acad Sci U S A. 2020. PMID: 31988119 Free PMC article.
-
A neuromechanical strategy for mediolateral foot placement in walking humans.J Neurophysiol. 2014 Jul 15;112(2):374-83. doi: 10.1152/jn.00138.2014. Epub 2014 Apr 30. J Neurophysiol. 2014. PMID: 24790168 Free PMC article.
References
-
- Stein BE, Stanford TR. Multisensory integration: current issues from the perspective of the single neuron. Nat Rev Neurosci. 2008;9:255–266. - PubMed
-
- Bulkin DA, Groh JM. Seeing sounds: visual and auditory interactions in the brain. Curr Opin Neurobiol. 2006;16:415–419. - PubMed
-
- Yuille AL, Bülthoff HH, editors. Bayesian decision theory and psychophysics. New York: Cambridge University Press; 1996.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
