

Neurexins physically and functionally interact with GABA(A) receptors
- PMID: 20471353
- PMCID: PMC3243752
- DOI: 10.1016/j.neuron.2010.04.008
Neurexins physically and functionally interact with GABA(A) receptors
Abstract
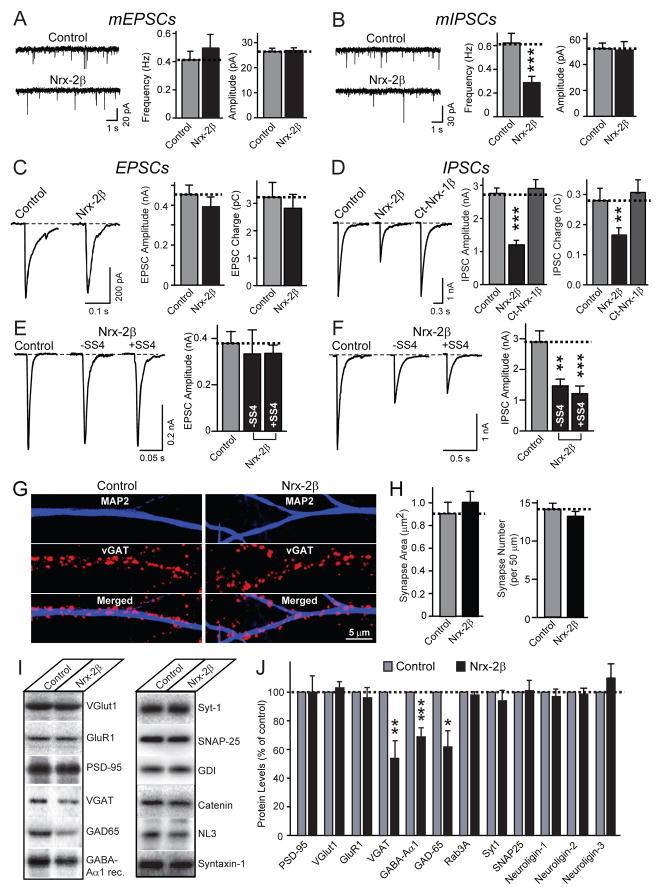
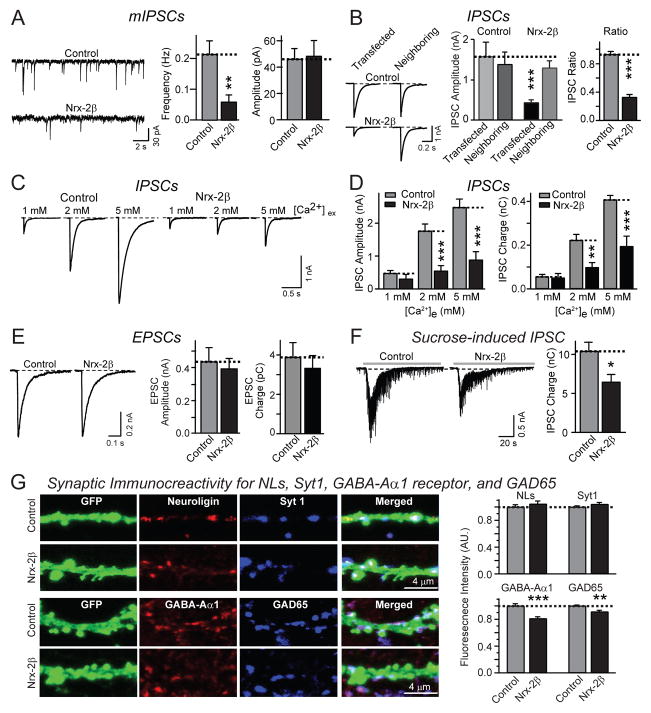
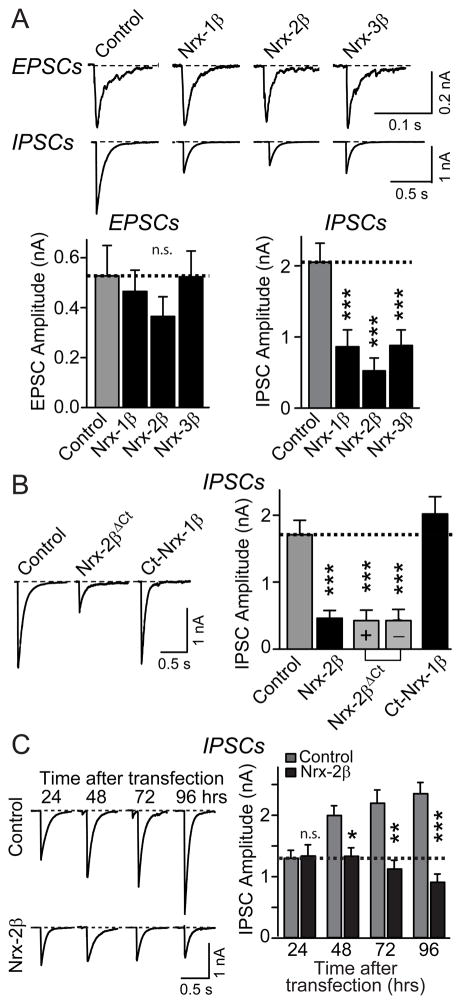
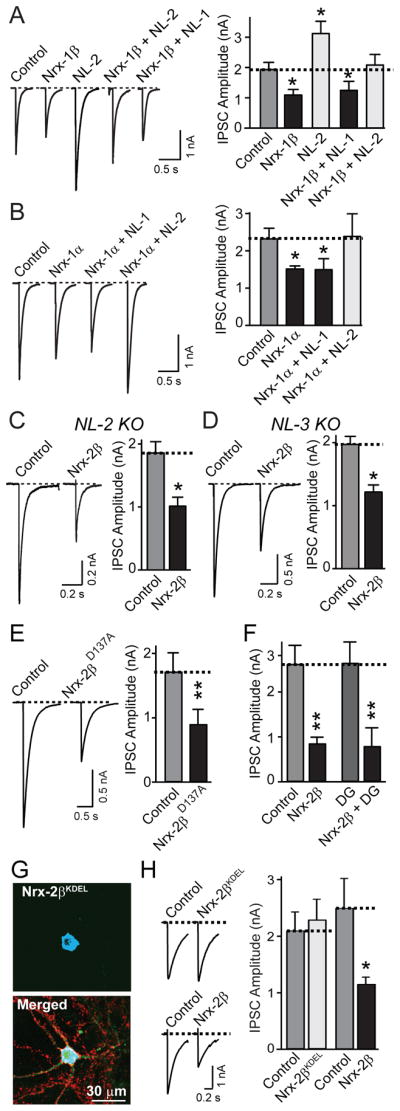
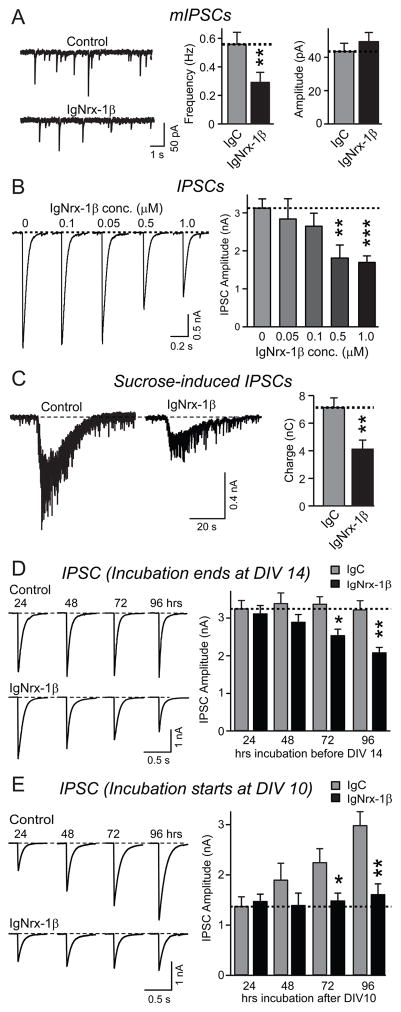
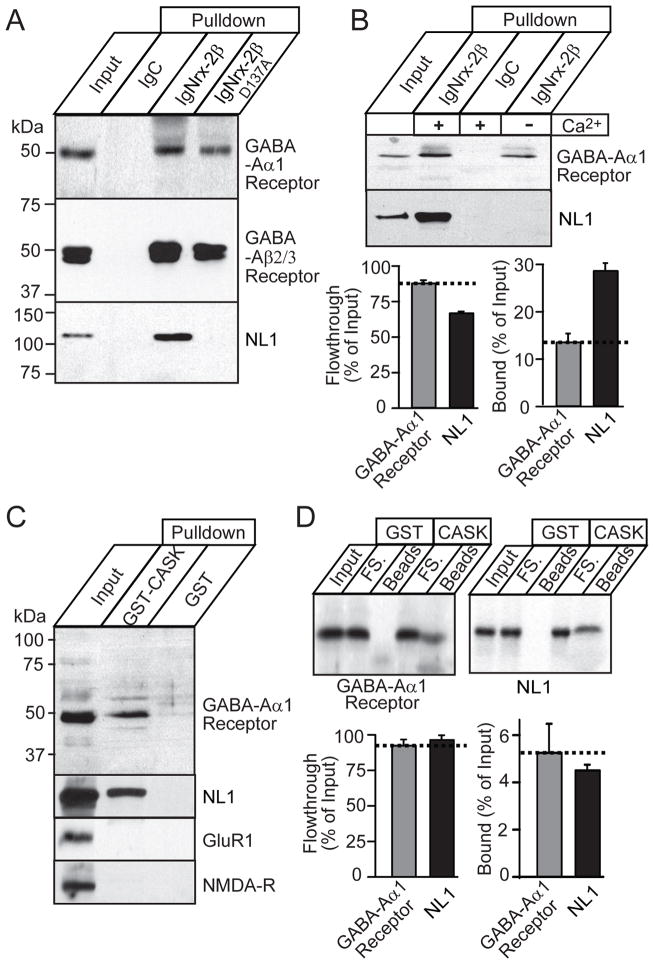
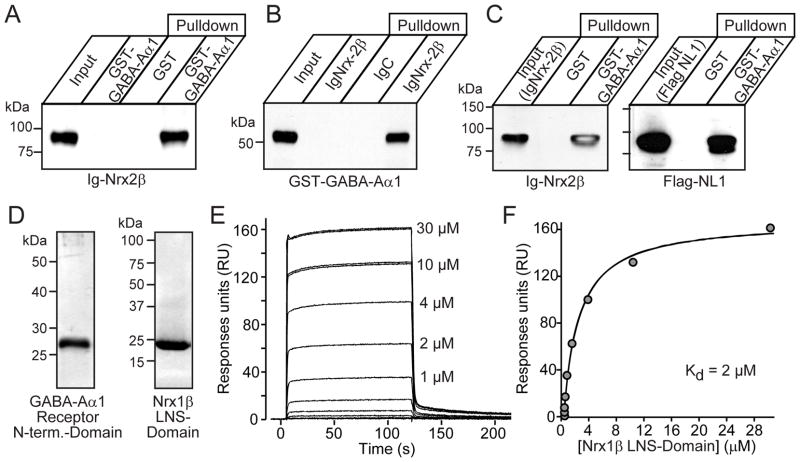
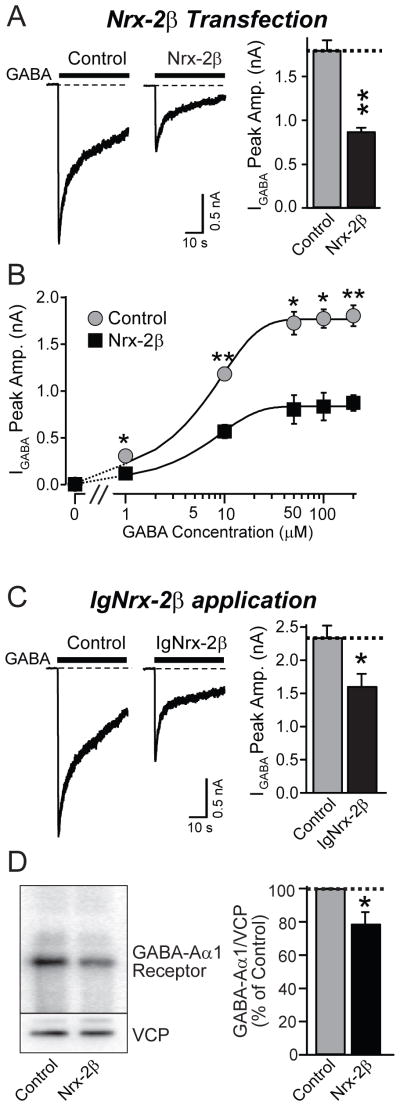
Neurexins are presynaptic cell-adhesion molecules that form trans-synaptic complexes with postsynaptic neuroligins. When overexpressed in nonneuronal cells, neurexins induce formation of postsynaptic specializations in cocultured neurons, suggesting that neurexins are synaptogenic. However, we find that when overexpressed in neurons, neurexins do not increase synapse density, but instead selectively suppressed GABAergic synaptic transmission without decreasing GABAergic synapse numbers. This suppression was mediated by all subtypes of neurexins tested, in a cell-autonomous and neuroligin-independent manner. Strikingly, addition of recombinant neurexin to cultured neurons at submicromolar concentrations induced the same suppression of GABAergic synaptic transmission as neurexin overexpression. Moreover, experiments with native brain proteins and purified recombinant proteins revealed that neurexins directly and stoichiometrically bind to GABA(A) receptors, suggesting that they decrease GABAergic synaptic responses by interacting with GABA(A) receptors. Our findings suggest that besides their other well-documented interactions, presynaptic neurexins directly act on postsynaptic GABA(A) receptors, which may contribute to regulate the excitatory/inhibitory balance in brain.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Araç D, Boucard AA, Özkan E, Strop P, Newell E, Südhof TC, Brunger AT. Structures of Neuroligin-1 and the Neuroligin-1/β-Neurexin-1 complex reveal specific protein-protein and protein-Ca2+ interactions. Neuron. 2007;56:992–1003. - PubMed
-
- Baumgartner S, Littleton JT, Broadie K, Bhat MA, Harbecke R, Lengyel JA, Chiquet-Ehrismann R, Prokop A, Bellen HJ. A Drosophila neurexin is required for septate junction and blood-nerve barrier formation and function. Cell. 1996;87:1059–1068. - PubMed
-
- Boucard AA, Chubykin AA, Comoletti D, Taylor P, Südhof TC. A splice code for trans-synaptic cell adhesion mediated by binding of neuroligin 1 to α- and β-neurexins. Neuron. 2005;48:229–236. - PubMed
-
- Chih B, Afridi SK, Clark L, Scheiffele P. Disorder-associated mutations lead to functional inactivation of neuroligins. Hum Mol Genet. 2004;13(14):1471–1477. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
