

Distinct contributions of rod, cone, and melanopsin photoreceptors to encoding irradiance
- PMID: 20471354
- PMCID: PMC2875410
- DOI: 10.1016/j.neuron.2010.04.037
Distinct contributions of rod, cone, and melanopsin photoreceptors to encoding irradiance
Abstract
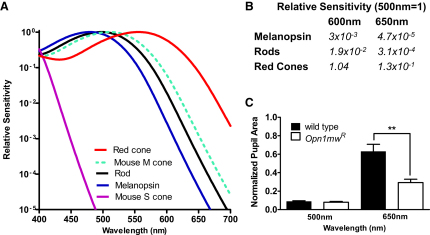
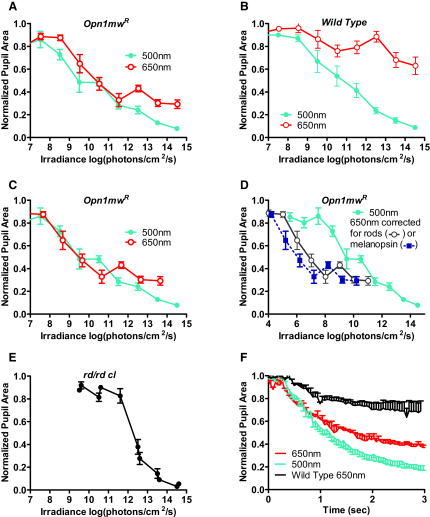
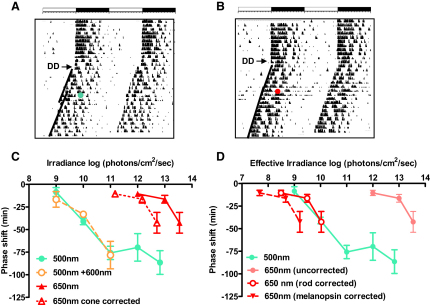
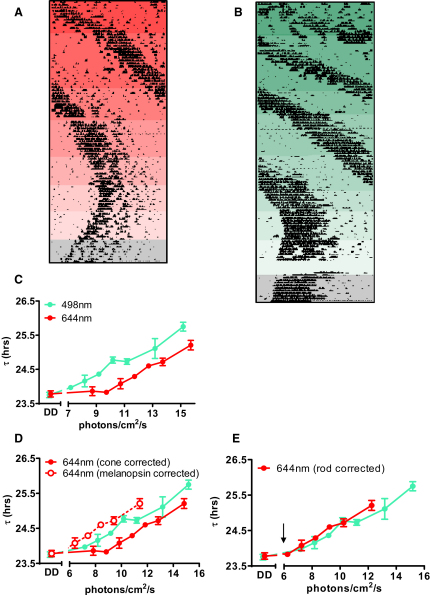
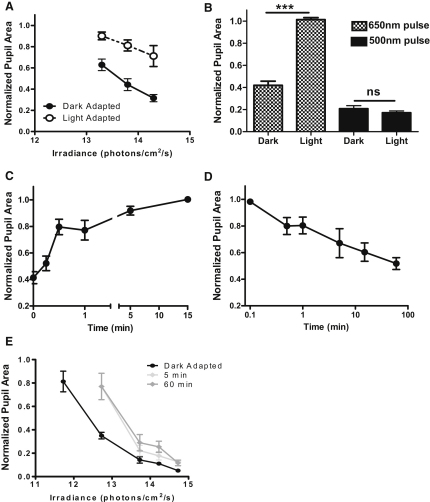
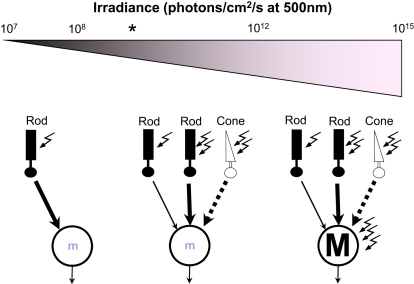
Photoreceptive, melanopsin-expressing retinal ganglion cells (mRGCs) encode ambient light (irradiance) for the circadian clock, the pupillomotor system, and other influential behavioral/physiological responses. mRGCs are activated both by their intrinsic phototransduction cascade and by the rods and cones. However, the individual contribution of each photoreceptor class to irradiance responses remains unclear. We address this deficit using mice expressing human red cone opsin, in which rod-, cone-, and melanopsin-dependent responses can be identified by their distinct spectral sensitivity. Our data reveal an unexpectedly important role for rods. These photoreceptors define circadian responses at very dim "scotopic" light levels but also at irradiances at which pattern vision relies heavily on cones. By contrast, cone input to irradiance responses dissipates following light adaptation to the extent that these receptors make a very limited contribution to circadian and pupillary light responses under these conditions. Our data provide new insight into retinal circuitry upstream of mRGCs and optimal stimuli for eliciting irradiance responses.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
