

Antiviral strategies to control calicivirus infections
- PMID: 20471996
- PMCID: PMC7114105
- DOI: 10.1016/j.antiviral.2010.05.002
Antiviral strategies to control calicivirus infections
Abstract
Caliciviridae are human or non-human pathogenic viruses with a high diversity. Some members of the Caliciviridae, i.e. human pathogenic norovirus or rabbit hemorrhagic disease virus (RHDV), are worldwide emerging pathogens. The norovirus is the major cause of viral gastroenteritis worldwide, accounting for about 85% of the outbreaks in Europe between 1995 and 2000. In the United States, 25 million cases of infection are reported each year. Since its emergence in 1984 as an agent of fatal hemorrhagic diseases in rabbits, RHDV has killed millions of rabbits and has been dispersed to all of the inhabitable continents. In view of their successful and apparently increasing emergence, the development of antiviral strategies to control infections due to these viral pathogens has now become an important issue in medicine and veterinary medicine. Antiviral strategies have to be based on an understanding of the epidemiology, transmission, clinical symptoms, viral replication and immunity to infection resulting from infection by these viruses. Here, we provide an overview of the mechanisms underlying calicivirus infection, focusing on the molecular aspects of replication in the host cell. Recent experimental data generated through an international collaboration on structural biology, virology and drug design within the European consortium VIZIER is also presented. Based on this analysis, we propose antiviral strategies that may significantly impact on the epidemiological characteristics of these highly successful viral pathogens.
Copyright 2010 Elsevier B.V. All rights reserved.
Figures
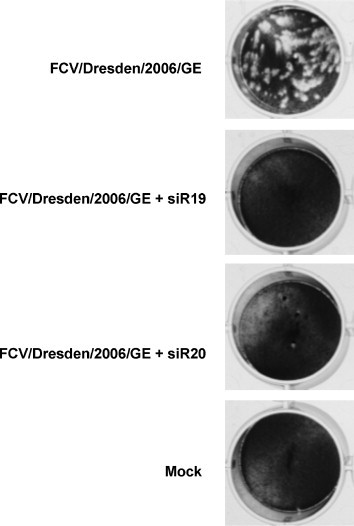
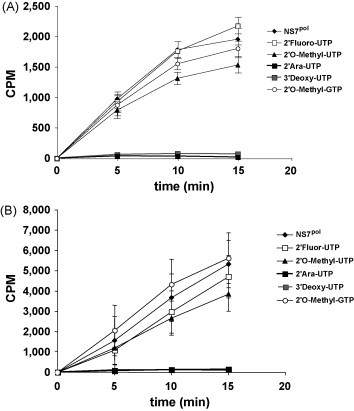
References
-
- Allaire M., Chernaia M.M., Malcolm B.A., James M.N. Picornaviral 3C cysteine proteinases have a fold similar to chymotrypsin-like serine proteinases. Nature. 1994;369:72–76. - PubMed
-
- Anand K., Ziebuhr J., Wadhwani P., Mesters J.R., Hilgenfeld R. Coronavirus main proteinase (3CLpro) structure: basis for design of anti-SARS drugs. Science. 2003;300:1763–1767. - PubMed
-
- Anderson E.J. Prevention and treatment of viral diarrhea in pediatrics. Expert Rev. Anti Infect. Ther. 2010;8:205–217. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
