

The genetic basis of laboratory adaptation in Caulobacter crescentus
- PMID: 20472802
- PMCID: PMC2897358
- DOI: 10.1128/JB.00255-10
The genetic basis of laboratory adaptation in Caulobacter crescentus
Abstract
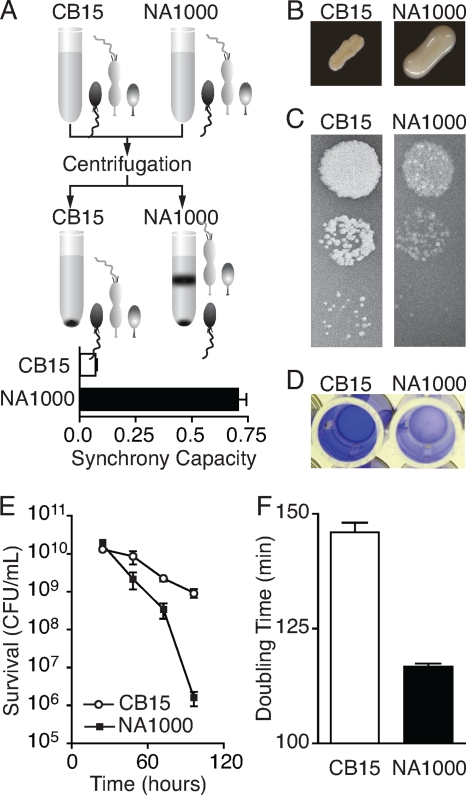
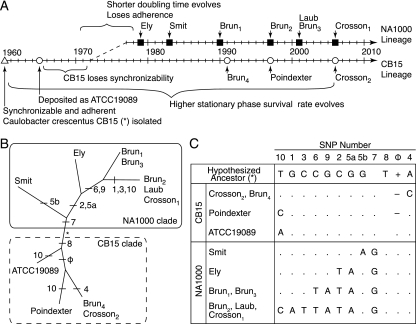
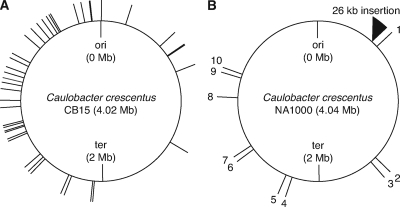
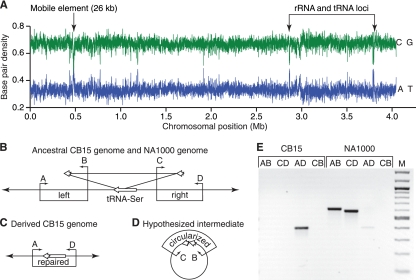
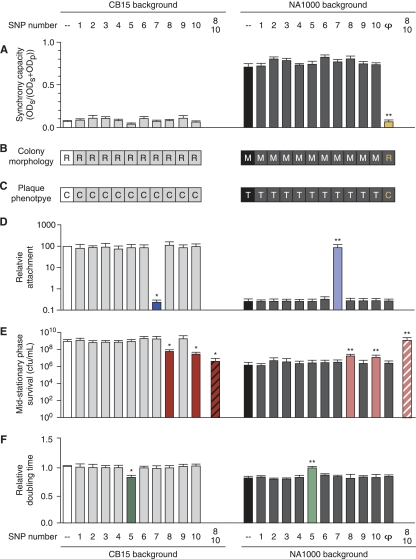
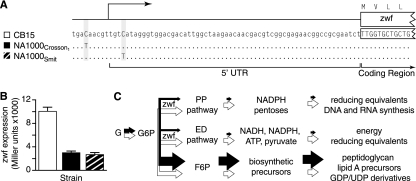
The dimorphic bacterium Caulobacter crescentus has evolved marked phenotypic changes during its 50-year history of culture in the laboratory environment, providing an excellent system for the study of natural selection and phenotypic microevolution in prokaryotes. Combining whole-genome sequencing with classical molecular genetic tools, we have comprehensively mapped a set of polymorphisms underlying multiple derived phenotypes, several of which arose independently in separate strain lineages. The genetic basis of phenotypic differences in growth rate, mucoidy, adhesion, sedimentation, phage susceptibility, and stationary-phase survival between C. crescentus strain CB15 and its derivative NA1000 is determined by coding, regulatory, and insertion/deletion polymorphisms at five chromosomal loci. This study evidences multiple genetic mechanisms of bacterial evolution as driven by selection for growth and survival in a new selective environment and identifies a common polymorphic locus, zwf, between lab-adapted C. crescentus and clinical isolates of Pseudomonas aeruginosa that have adapted to a human host during chronic infection.
Figures
References
-
- Artamonova, V. S., and A. A. Makhrov. 2006. Unintentional genetic processes in artificially maintained populations: proving the leading role of selection in evolution. Russ. J. Genet. 42:234-246. - PubMed
-
- Barrick, J. E., D. S. Yu, S. H. Yoon, H. Jeong, T. K. Oh, D. Schneider, R. E. Lenski, and J. F. Kim. 2009. Genome evolution and adaptation in a long-term experiment with Escherichia coli. Nature 461:1243-1247. - PubMed
-
- Boras, J. A., M. M. Sala, E. Vazquez-Dominguez, M. G. Weinbauer, and D. Vaque. 2009. Annual changes of bacterial mortality due to viruses and protists in an oligotrophic coastal environment (NW Mediterranean). Environ. Microbiol. 11:1181-1193. - PubMed
-
- Buckling, A., R. C. Maclean, M. A. Brockhurst, and N. Colegrave. 2009. The beagle in a bottle. Nature 457:824-829. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
