

cis-Regulatory elements and chromatin state coordinately control temporal and spatial expression of FLOWERING LOCUS T in Arabidopsis
- PMID: 20472817
- PMCID: PMC2899882
- DOI: 10.1105/tpc.110.074682
cis-Regulatory elements and chromatin state coordinately control temporal and spatial expression of FLOWERING LOCUS T in Arabidopsis
Abstract
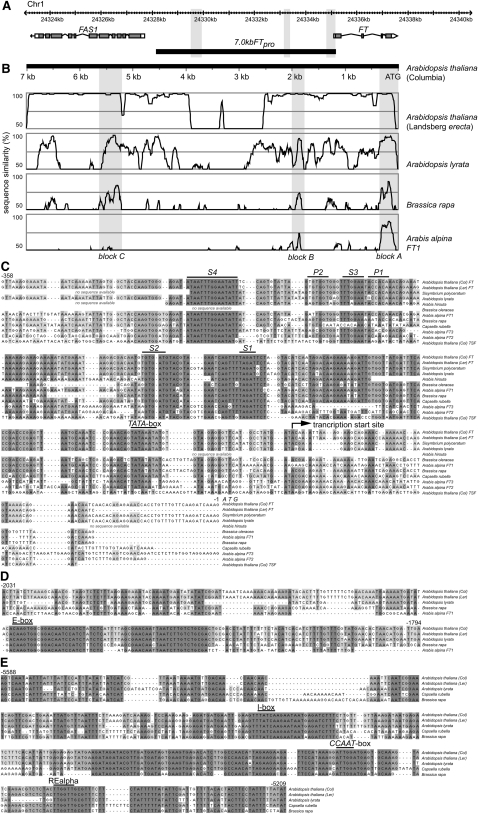
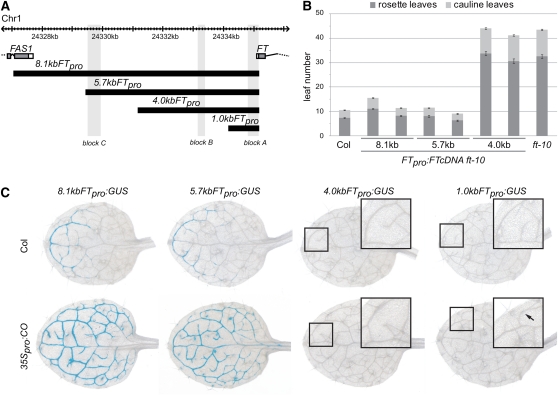
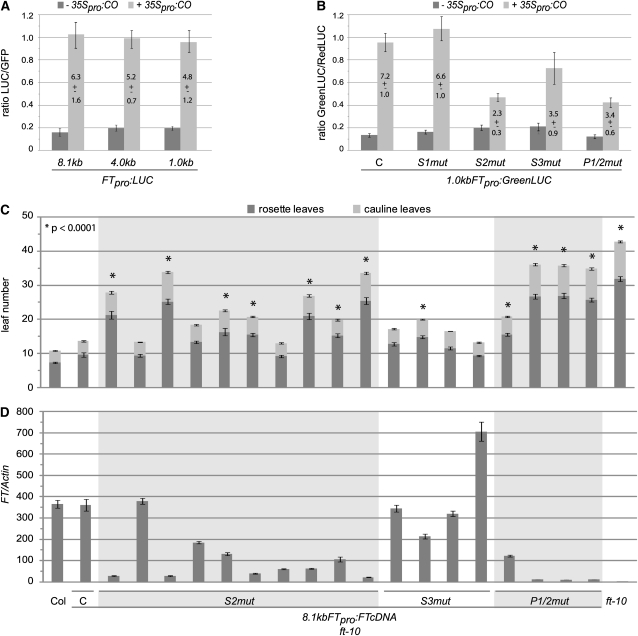
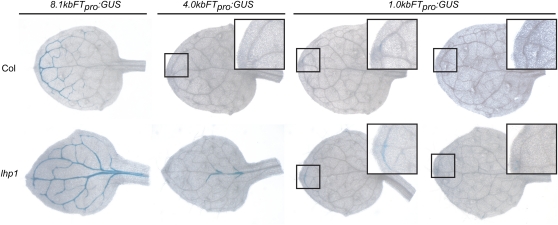
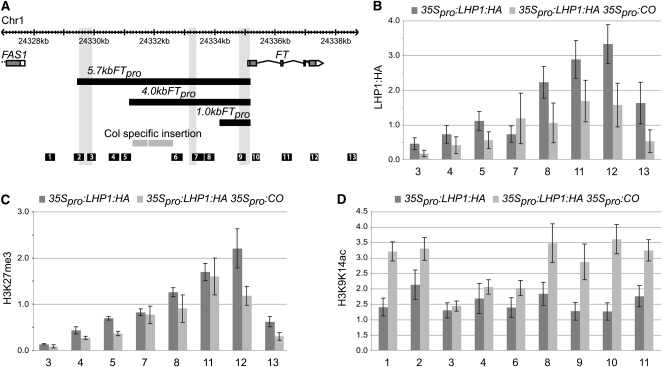
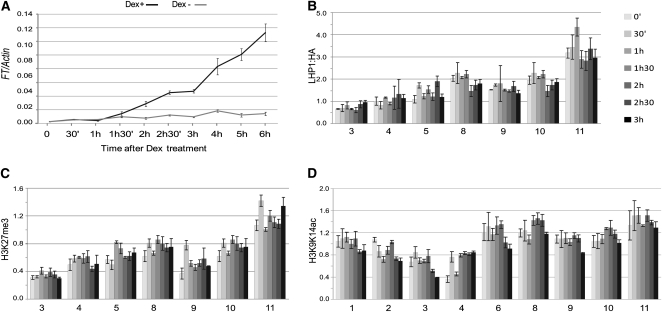
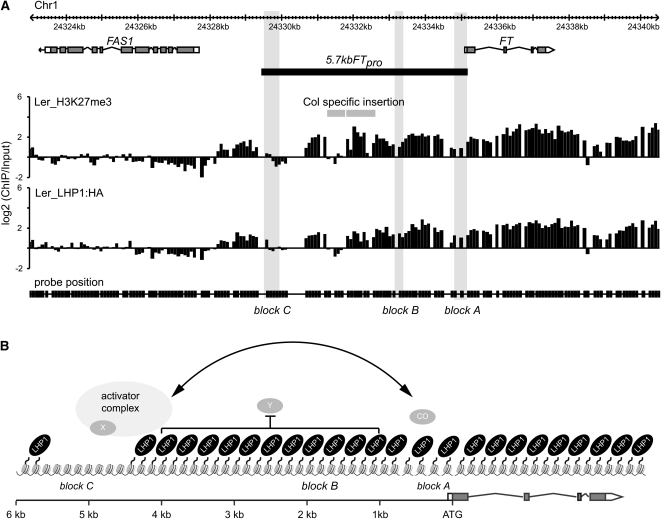
Flowering time of summer annual Arabidopsis thaliana accessions is largely determined by the timing of FLOWERING LOCUS T (FT) expression in the leaf vasculature. To understand the complex interplay between activating and repressive inputs controlling flowering through FT, cis-regulatory sequences of FT were identified in this study. A proximal and an approximately 5-kb upstream promoter region containing highly conserved sequence blocks were found to be essential for FT activation by CONSTANS (CO). Chromatin-associated protein complexes add another layer to FT regulation. In plants constitutively overexpressing CO, changes in chromatin status, such as a decrease in binding of LIKE HETEROCHROMATIN PROTEIN1 (LHP1) and increased acetylation of H3K9 and K14, were observed throughout the FT locus, although these changes appear to be a consequence of FT upregulation and not a prerequisite for activation. Binding of LHP1 was required to repress enhancer elements located between the CO-controlled regions. By contrast, the distal and proximal promoter sequences required for FT activation coincide with locally LHP1 and H3K27me3 depleted chromatin, indicating that chromatin status facilitates the accessibility of transcription factors to FT. Therefore, distant regulatory regions are required for FT transcription, reflecting the complexity of its control and differences in chromatin status delimit functionally important cis-regulatory regions.
Figures
Comment in
-
Dissecting cis-regulation of FLOWERING LOCUS T.Plant Cell. 2010 May;22(5):1422. doi: 10.1105/tpc.110.220511. Epub 2010 May 18. Plant Cell. 2010. PMID: 20484006 Free PMC article. No abstract available.
References
-
- Abe M., Kobayashi Y., Yamamoto S., Daimon Y., Yamaguchi A., Ikeda Y., Ichinoki H., Notaguchi M., Goto K., Araki T. (2005). FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309: 1052–1056 - PubMed
-
- Ben-Naim O., Parnis R.E.A., Teper-Bamnolker P., Shalit A., Coupland G., Samach A., Lifschitz E. (2006). The CCAAT binding factor can mediate interactions between CONSTANS-like proteins and DNA. Plant J. 46: 462–476 - PubMed
-
- Bradley D., Ratcliffe O., Vincent C., Carpenter R., Coen E. (1997). Inflorescence commitment and architecture in Arabidopsis. Science 275: 80–83 - PubMed
-
- Brudno M., Malde S., Poliakov A., Do C.B., Couronne O., Dubchak I., Batzoglou S. (2003). Glocal alignment: Finding rearrangements during alignment. Bioinformatics 19: i54–i62 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
