

Actin-depolymerizing factor cofilin-1 is necessary in maintaining mature podocyte architecture
- PMID: 20472933
- PMCID: PMC2903407
- DOI: 10.1074/jbc.M110.122929
Actin-depolymerizing factor cofilin-1 is necessary in maintaining mature podocyte architecture
Abstract
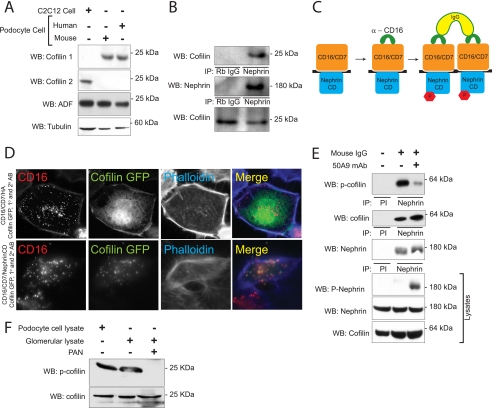
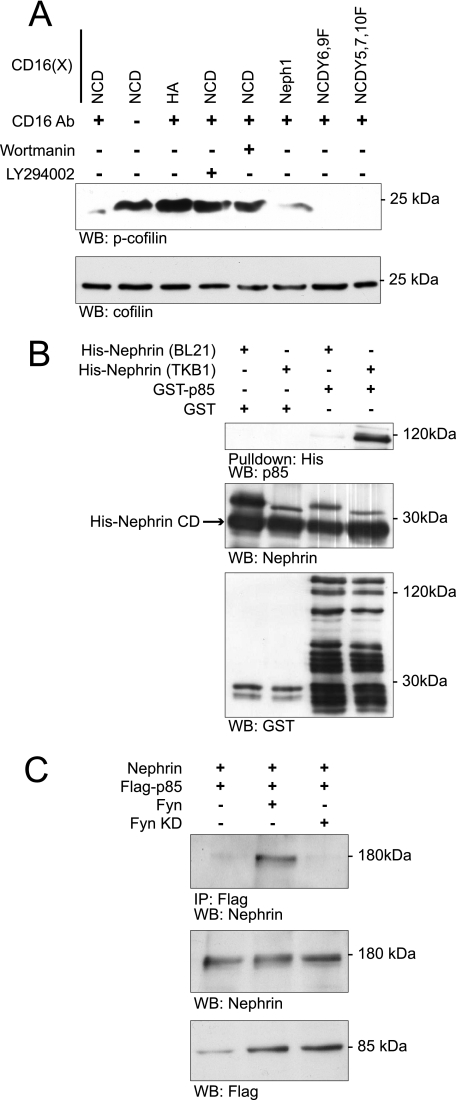
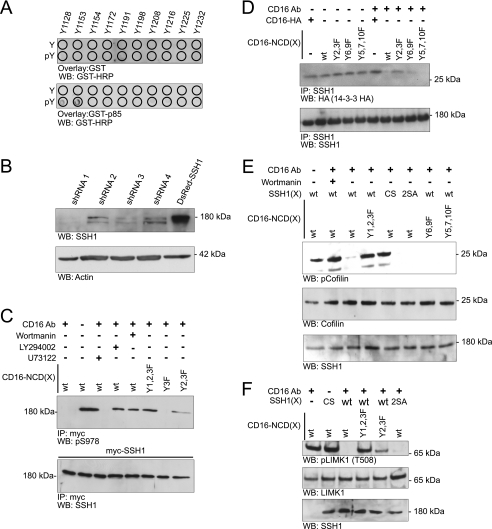
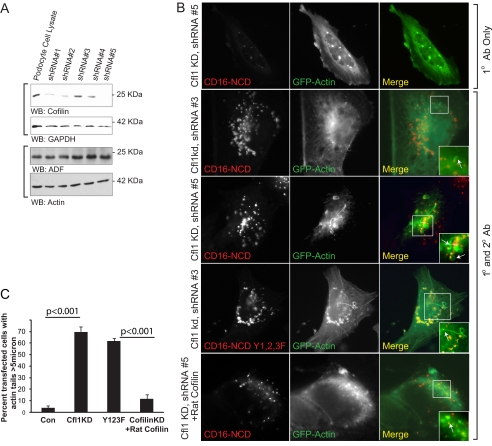
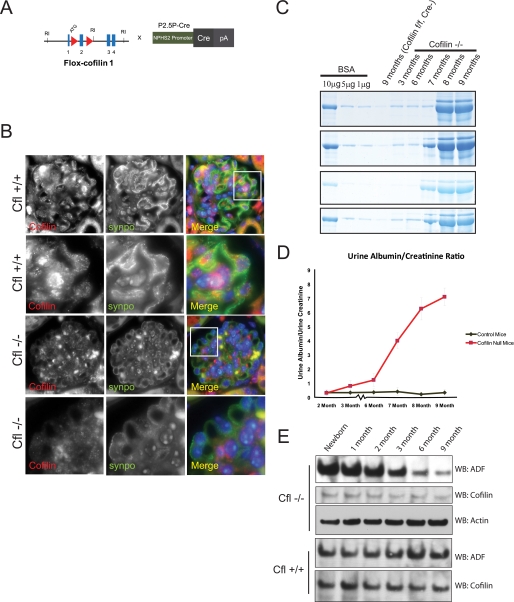
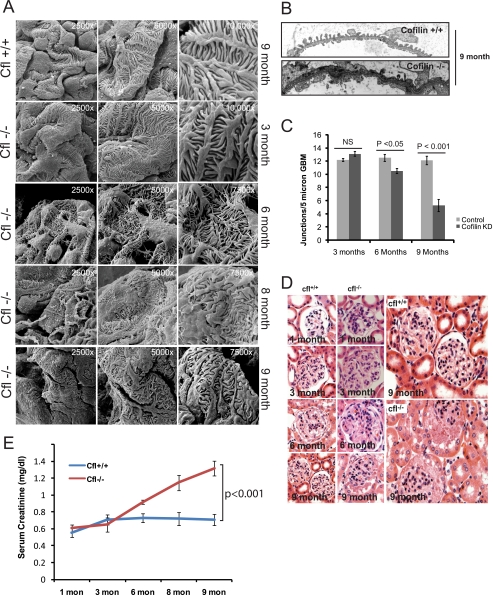
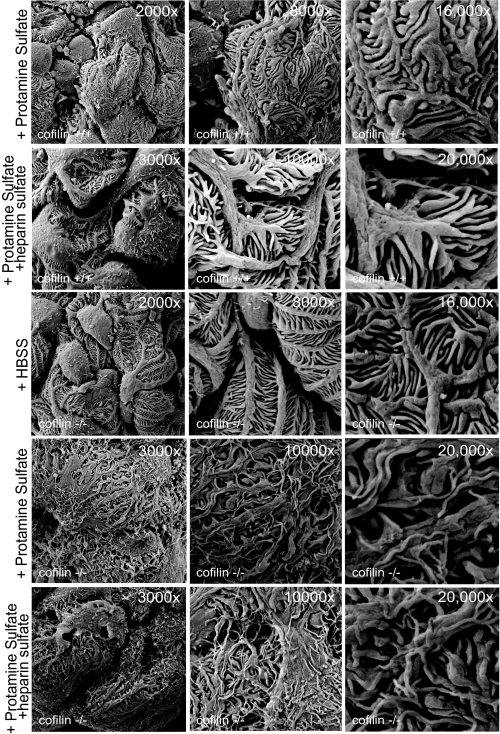
Actin dynamics determines podocyte morphology during development and in response to podocyte injury and might be necessary for maintaining normal podocyte morphology. Because podocyte intercellular junction receptor Nephrin plays a role in regulating actin dynamics, and given the described role of cofilin in actin filament polymerization and severing, we hypothesized that cofilin-1 activity is regulated by Nephrin and is necessary in normal podocyte actin dynamics. Nephrin activation induced cofilin dephosphorylation via intermediaries that include phosphatidylinositol 3-kinase, SSH1, 14-3-3, and LIMK in a cell culture model. This Nephrin-induced cofilin activation required a direct interaction between Nephrin and the p85 subunit of phosphatidylinositol 3-kinase. In a similar fashion, cofilin-1 dephosphorylation was observed in a rat model of podocyte injury at a time when foot process spreading is initially observed. To investigate the necessity of cofilin-1 in the glomerulus, podocyte-specific Cfl1 null mice were generated. Cfl1 null podocytes developed normally. However, these mice developed persistent proteinuria by 3 months of age, although they did not exhibit foot process spreading until 8 months, when the rate of urinary protein excretion became more exaggerated. In a mouse model of podocyte injury, protamine sulfate perfusion of the Cfl1 mutant mouse induced a broadened and flattened foot process morphology that was distinct from that observed following perfusion of control kidneys, and mutant podocytes did not recover normal structure following additional perfusion with heparin sulfate. We conclude that cofilin-1 is necessary for maintenance of normal podocyte architecture and for actin structural changes that occur during induction and recovery from podocyte injury.
Figures
References
-
- Kestilä M., Lenkkeri U., Männikkö M., Lamerdin J., McCready P., Putaala H., Ruotsalainen V., Morita T., Nissinen M., Herva R., Kashtan C. E., Peltonen L., Holmberg C., Olsen A., Tryggvason K. (1998) Mol. Cell 1, 575–582 - PubMed
-
- Kim J. M., Wu H., Green G., Winkler C. A., Kopp J. B., Miner J. H., Unanue E. R., Shaw A. S. (2003) Science 300, 1298–1300 - PubMed
-
- Winn M. P., Conlon P. J., Lynn K. L., Farrington M. K., Creazzo T., Hawkins A. F., Daskalakis N., Kwan S. Y., Ebersviller S., Burchette J. L., Pericak-Vance M. A., Howell D. N., Vance J. M., Rosenberg P. B. (2005) Science 308, 1801–1804 - PubMed
-
- Kaplan J. M., Kim S. H., North K. N., Rennke H., Correia L. A., Tong H. Q., Mathis B. J., Rodríguez-Pérez J. C., Allen P. G., Beggs A. H., Pollak M. R. (2000) Nat. Genet. 24, 251–256 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
