

A'-form RNA helices are required for cytoplasmic mRNA transport in Drosophila
- PMID: 20473315
- PMCID: PMC2997434
- DOI: 10.1038/nsmb.1813
A'-form RNA helices are required for cytoplasmic mRNA transport in Drosophila
Abstract
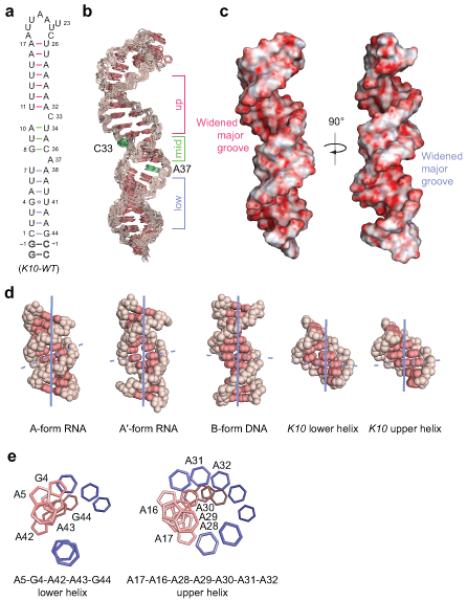
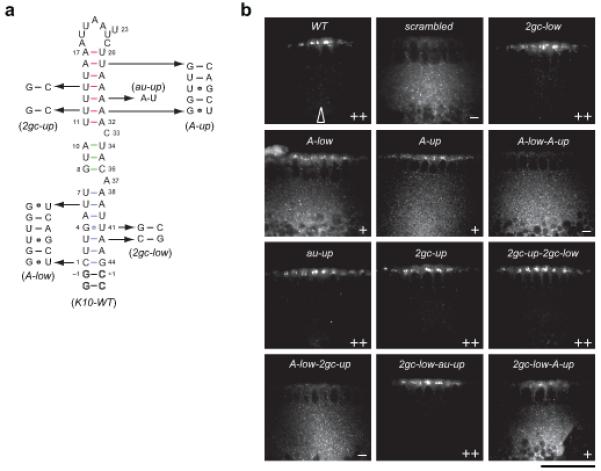
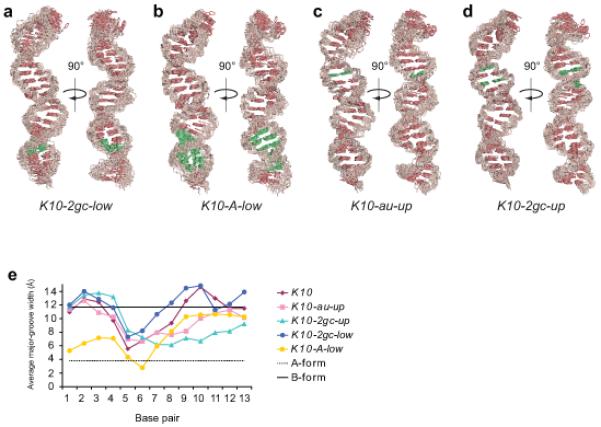
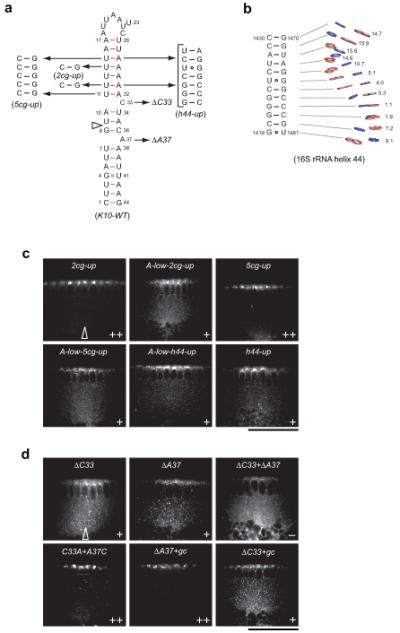
Microtubule-based mRNA transport is widely used to restrict protein expression to specific regions in the cell and has important roles in defining cell polarity and axis determination as well as in neuronal function. However, the structural basis of recognition of cis-acting mRNA localization signals by motor complexes is poorly understood. We have used NMR spectroscopy to describe the first tertiary structure to our knowledge of an RNA element responsible for mRNA transport. The Drosophila melanogaster fs(1)K10 signal, which mediates transport by the dynein motor, forms a stem loop with two double-stranded RNA helices adopting an unusual A'-form conformation with widened major grooves reminiscent of those in B-form DNA. Structure determination of four mutant RNAs and extensive functional assays in Drosophila embryos indicate that the two spatially registered A'-form helices represent critical recognition sites for the transport machinery. Our study provides insights into the basis for RNA cargo recognition and reveals a key biological function encoded by A'-form RNA conformation.
Figures
References
-
- Wilkie GS, Davis I. Drosophila wingless and pair-rule transcripts localize apically by dynein-mediated transport of RNA particles. Cell. 2001;105:209–19. - PubMed
-
- Van De Bor V, Hartswood E, Jones C, Finnegan D, Davis I. gurken and the I factor retrotransposon RNAs share common localization signals and machinery. Dev Cell. 2005;9:51–62. - PubMed
References for online methods only
-
- Nikonowicz EP. Preparation and use of 2H-labeled RNA oligonucleotides in nuclear magnetic resonance studies. Methods Enzymol. 2001;338:320–41. - PubMed
-
- Scott LG, Tolbert TJ, Williamson JR. Preparation of specifically 2H- and 13C-labeled ribonucleotides. Methods Enzymol. 2000;317:18–38. - PubMed
-
- Hansen MR, Hanson P, Pardi A. Filamentous bacteriophage for aligning RNA, DNA, and proteins for measurement of nuclear magnetic resonance dipolar coupling interactions. Methods Enzymol. 2000;317:220–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
