

Design of Mn porphyrins for treating oxidative stress injuries and their redox-based regulation of cellular transcriptional activities
- PMID: 20473774
- PMCID: PMC3022969
- DOI: 10.1007/s00726-010-0603-6
Design of Mn porphyrins for treating oxidative stress injuries and their redox-based regulation of cellular transcriptional activities
Erratum in
- Amino Acids. 2012 Jan;42(1):115-6
Abstract
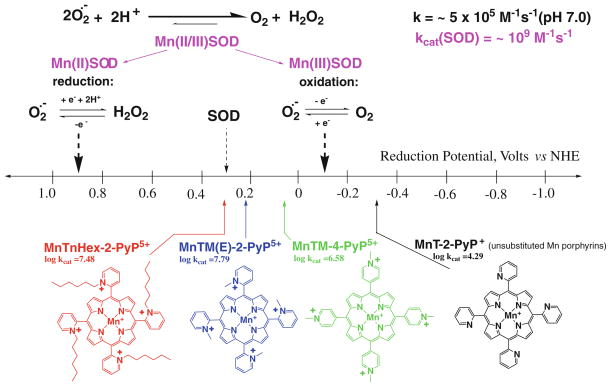
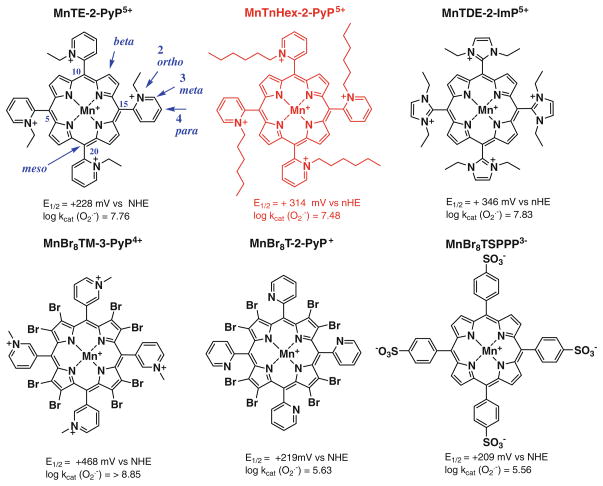
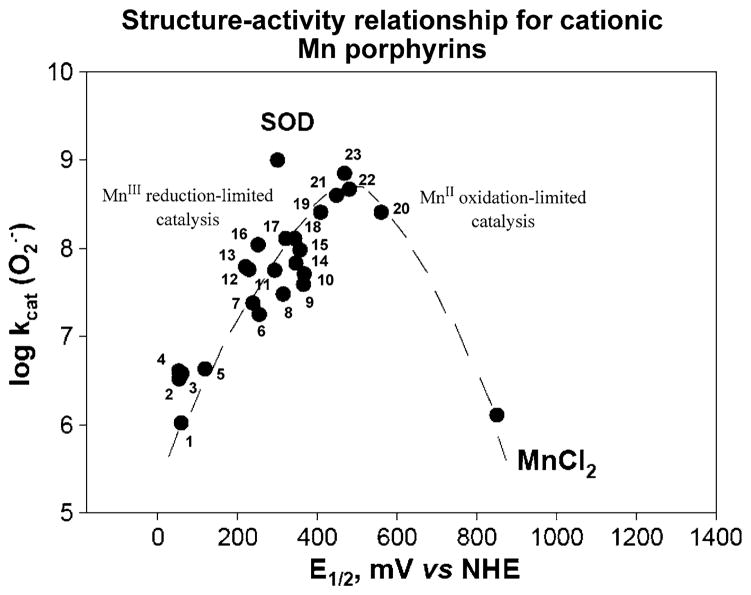
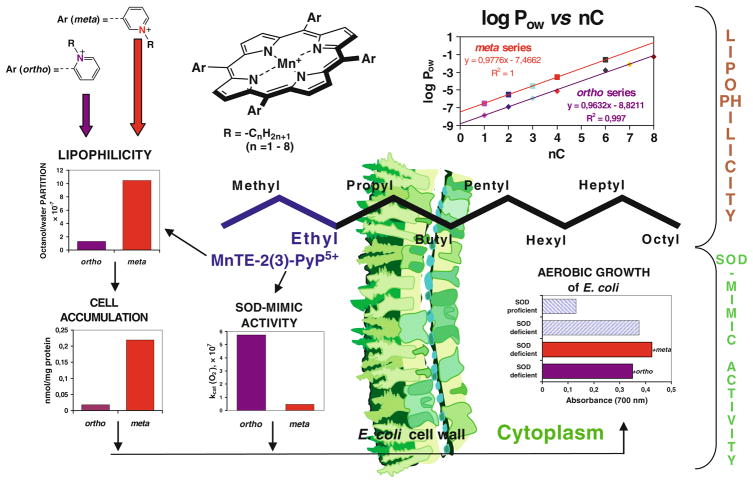
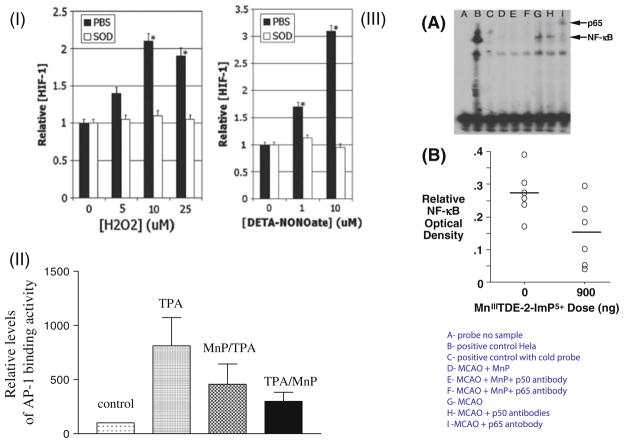
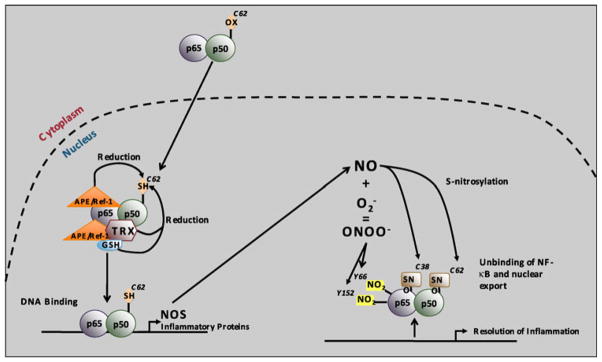
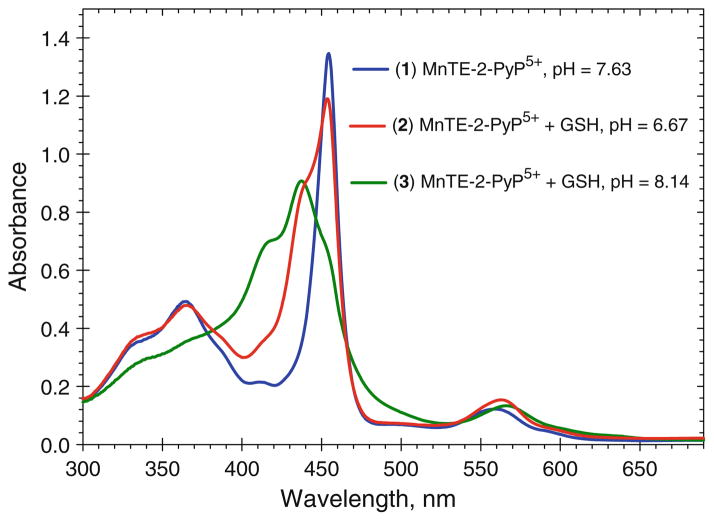
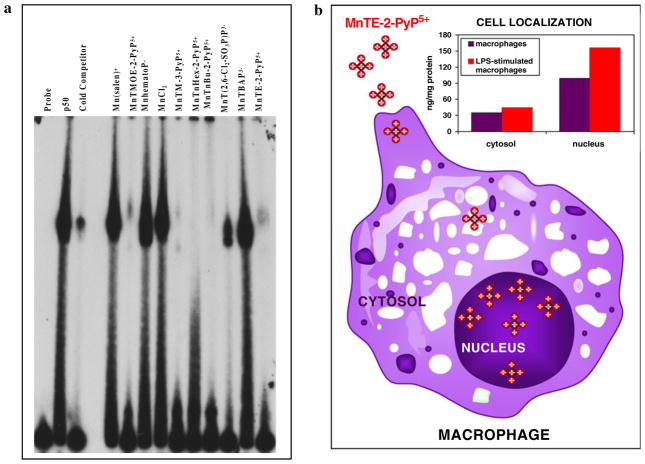
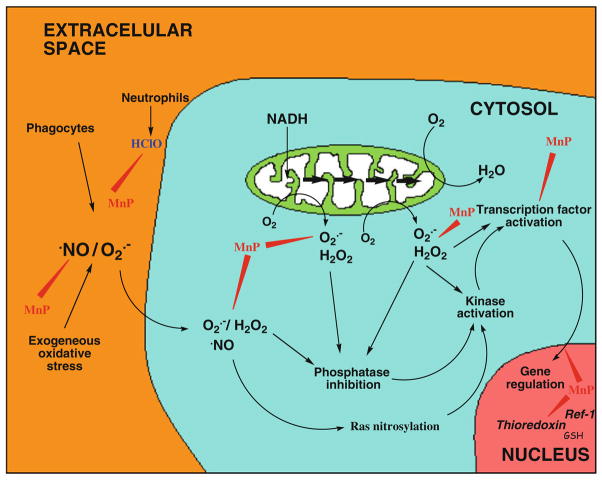
The most efficacious Mn(III) porphyrinic (MnPs) scavengers of reactive species have positive charges close to the Mn site, whereby they afford thermodynamic and electrostatic facilitation for the reaction with negatively charged species such as O (2) (•-) and ONOO(-). Those are Mn(III) meso tetrakis(N-alkylpyridinium-2-yl)porphyrins, more specifically MnTE-2-PyP(5+) (AEOL10113) and MnTnHex-2-PyP(5+) (where alkyls are ethyl and n-hexyl, respectively), and their imidazolium analog, MnTDE-2-ImP(5+) (AEOL10150, Mn(III) meso tetrakis(N,N'-diethylimidazolium-2-yl) porphyrin). The efficacy of MnPs in vivo is determined not only by the compound antioxidant potency, but also by its bioavailability. The former is greatly affected by the lipophilicity, size, structure, and overall shape of the compound. These porphyrins have the ability to both eliminate reactive oxygen species and impact the progression of oxidative stress-dependent signaling events. This will effectively lead to the regulation of redox-dependent transcription factors and the suppression of secondary inflammatory- and oxidative stress-mediated immune responses. We have reported on the inhibition of major transcription factors HIF-1α, AP-1, SP-1, and NF-κB by Mn porphyrins. While the prevailing mechanistic view of the suppression of transcription factors activation is via antioxidative action (presumably in cytosol), the pro-oxidative action of MnPs in suppressing NF-κB activation in nucleus has been substantiated. The magnitude of the effect is dependent upon the electrostatic (porphyrin charges) and thermodynamic factors (porphyrin redox ability). The pro-oxidative action of MnPs has been suggested to contribute at least in part to the in vitro anticancer action of MnTE-2-PyP(5+) in the presence of ascorbate, and in vivo when combined with chemotherapy of lymphoma. Given the remarkable therapeutic potential of metalloporphyrins, future studies are warranted to further our understanding of in vivo action/s of Mn porphyrins, particularly with respect to their subcellular distribution.
Figures
References
-
- Al-Mutairi D, Craik JD, Batinic-Haberle I, Benov L. Inactivation of metabolic enzymes by photo-treatment with zinc meta N-methylpyridylporphyrin. Biochim Biophys Acta. 2007;1770:1520–1527. - PubMed
-
- Archibald FS, Fridovich I. The scavenging of superoxide radical by manganous complexes: in vitro. Arch Biochem Biophys. 1982;214:452–463. - PubMed
-
- Barnese K, Gralla EB, Cabelli DE, Valentine JS. Manganous phosphate acts as a superoxide dismutase. J Am Chem Soc. 2008;130:4604–4606. - PubMed
-
- Batinic-Haberle I, Benov L, Spasojević I, Fridovich I. The ortho effect makes manganese (III) meso-tetrakis(N-methylpy-ridinium-2-yl)porphyrin (MnTM-2-PyP) a powerful and potentially useful superoxide dismutase mimic. J Biol Chem. 1998;273:24521–24528. - PubMed
-
- Batinic-Haberle I, Benov L, Spasojević I, Hambright P, Crumbliss AL, Fridovich I. The relationship between redox potentials, proton dissociation constants of pyrrolic nitrogens, and in vitro and in vivo superoxide dismutase activities of manganese(III) and iron(III) cationic and anionic porphyrins. Inorg Chem. 1999;38:4011–4022.
Publication types
MeSH terms
Substances
Grants and funding
- P01 CA042745/CA/NCI NIH HHS/United States
- CA40355-25/CA/NCI NIH HHS/United States
- 5-P30-CA14236-29/CA/NCI NIH HHS/United States
- R01 DA024074/DA/NIDA NIH HHS/United States
- R01 CA139843/CA/NCI NIH HHS/United States
- U19AI067798/AI/NIAID NIH HHS/United States
- U19 AI067798/AI/NIAID NIH HHS/United States
- R01 CA098452/CA/NCI NIH HHS/United States
- P30 CA014236/CA/NCI NIH HHS/United States
- CA 07359/CA/NCI NIH HHS/United States
- 1 UL 1 RR024128-01/RR/NCRR NIH HHS/United States
- CA 139843/CA/NCI NIH HHS/United States
- R01 CA 098452/CA/NCI NIH HHS/United States
- R01 CA040355/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
